

The functional role of sequentially neuromodulated synaptic plasticity in behavioural learning
- PMID: 34111110
- PMCID: PMC8192019
- DOI: 10.1371/journal.pcbi.1009017
The functional role of sequentially neuromodulated synaptic plasticity in behavioural learning
Abstract
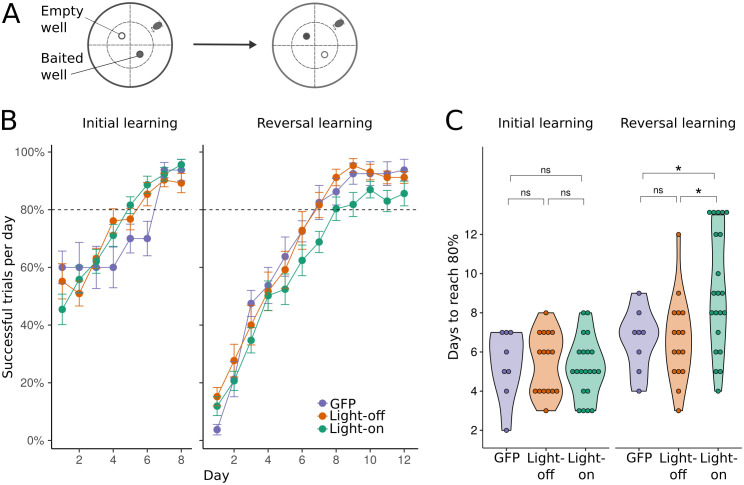
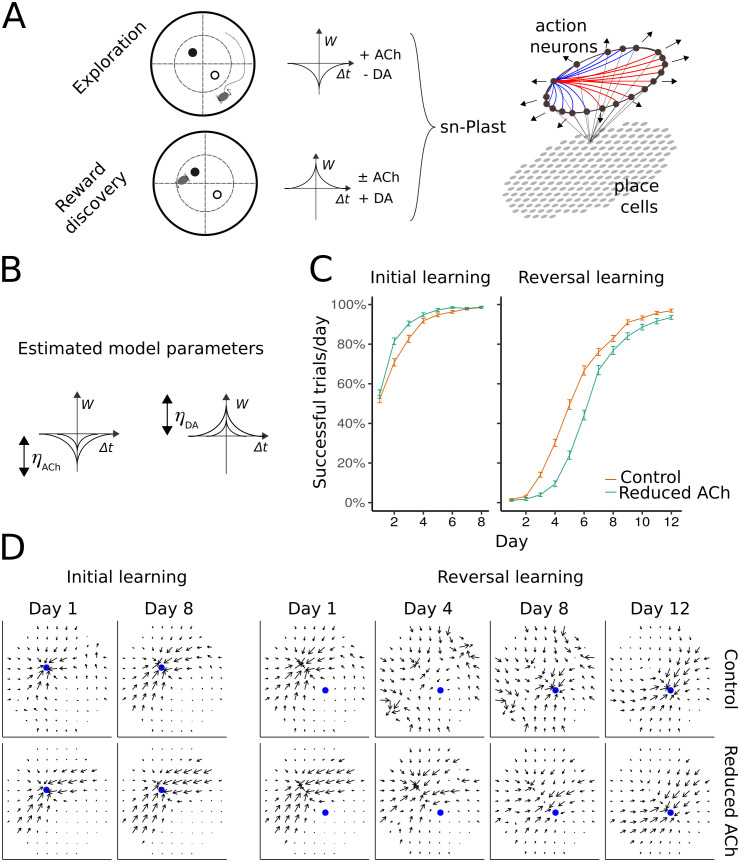
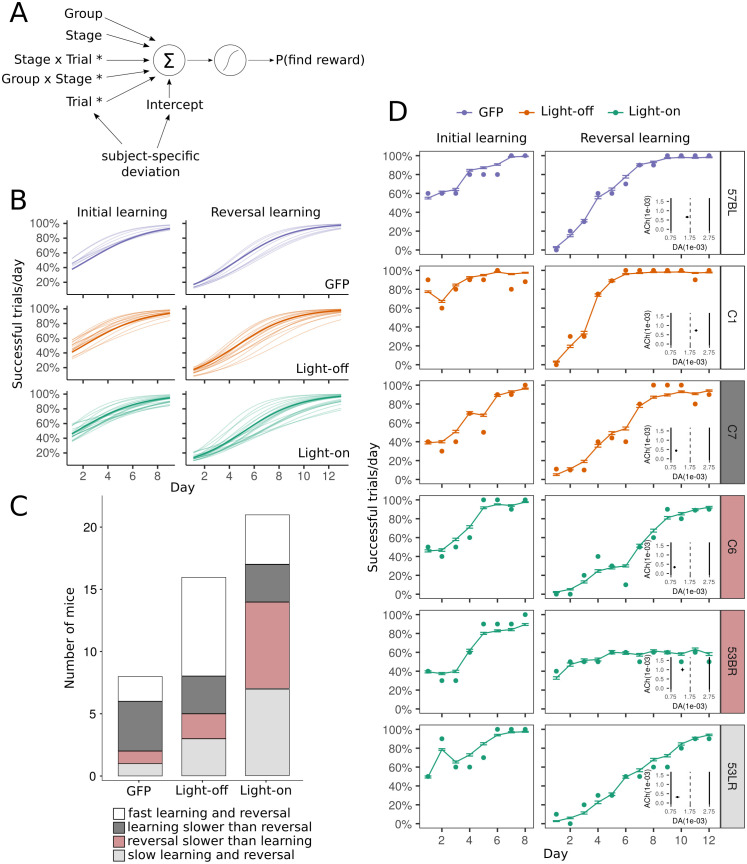
To survive, animals have to quickly modify their behaviour when the reward changes. The internal representations responsible for this are updated through synaptic weight changes, mediated by certain neuromodulators conveying feedback from the environment. In previous experiments, we discovered a form of hippocampal Spike-Timing-Dependent-Plasticity (STDP) that is sequentially modulated by acetylcholine and dopamine. Acetylcholine facilitates synaptic depression, while dopamine retroactively converts the depression into potentiation. When these experimental findings were implemented as a learning rule in a computational model, our simulations showed that cholinergic-facilitated depression is important for reversal learning. In the present study, we tested the model's prediction by optogenetically inactivating cholinergic neurons in mice during a hippocampus-dependent spatial learning task with changing rewards. We found that reversal learning, but not initial place learning, was impaired, verifying our computational prediction that acetylcholine-modulated plasticity promotes the unlearning of old reward locations. Further, differences in neuromodulator concentrations in the model captured mouse-by-mouse performance variability in the optogenetic experiments. Our line of work sheds light on how neuromodulators enable the learning of new contingencies.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Acetylcholine-modulated plasticity in reward-driven navigation: a computational study.Sci Rep. 2018 Jun 21;8(1):9486. doi: 10.1038/s41598-018-27393-2. Sci Rep. 2018. PMID: 29930322 Free PMC article.
-
Sequential neuromodulation of Hebbian plasticity offers mechanism for effective reward-based navigation.Elife. 2017 Jul 10;6:e27756. doi: 10.7554/eLife.27756. Elife. 2017. PMID: 28691903 Free PMC article.
-
Functional requirements for reward-modulated spike-timing-dependent plasticity.J Neurosci. 2010 Oct 6;30(40):13326-37. doi: 10.1523/JNEUROSCI.6249-09.2010. J Neurosci. 2010. PMID: 20926659 Free PMC article.
-
Neuromodulated Spike-Timing-Dependent Plasticity, and Theory of Three-Factor Learning Rules.Front Neural Circuits. 2016 Jan 19;9:85. doi: 10.3389/fncir.2015.00085. eCollection 2015. Front Neural Circuits. 2016. PMID: 26834568 Free PMC article. Review.
-
Neuromodulation of Spike-Timing-Dependent Plasticity: Past, Present, and Future.Neuron. 2019 Aug 21;103(4):563-581. doi: 10.1016/j.neuron.2019.05.041. Neuron. 2019. PMID: 31437453 Review.
Cited by
-
A Navigation Path Search and Optimization Method for Mobile Robots Based on the Rat Brain's Cognitive Mechanism.Biomimetics (Basel). 2023 Sep 14;8(5):427. doi: 10.3390/biomimetics8050427. Biomimetics (Basel). 2023. PMID: 37754178 Free PMC article.
-
[Spatial navigation method based on the entorhinal-hippocampal-prefrontal information transmission circuit of rat's brain].Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 2024 Feb 25;41(1):80-89. doi: 10.7507/1001-5515.202303047. Sheng Wu Yi Xue Gong Cheng Xue Za Zhi. 2024. PMID: 38403607 Free PMC article. Chinese.
References
Publication types
MeSH terms
Substances
Grants and funding
- BB/P019560/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 200790/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- BB/N019008/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- BB/N013956/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
