

Replication protein A plays multifaceted roles complementary to specialized helicases in processing G-quadruplex DNA
- PMID: 34113828
- PMCID: PMC8169993
- DOI: 10.1016/j.isci.2021.102493
Replication protein A plays multifaceted roles complementary to specialized helicases in processing G-quadruplex DNA
Abstract
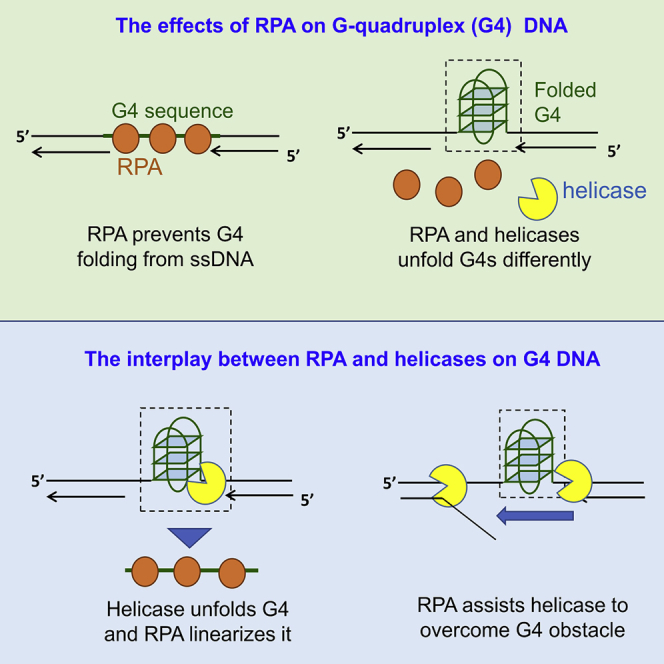
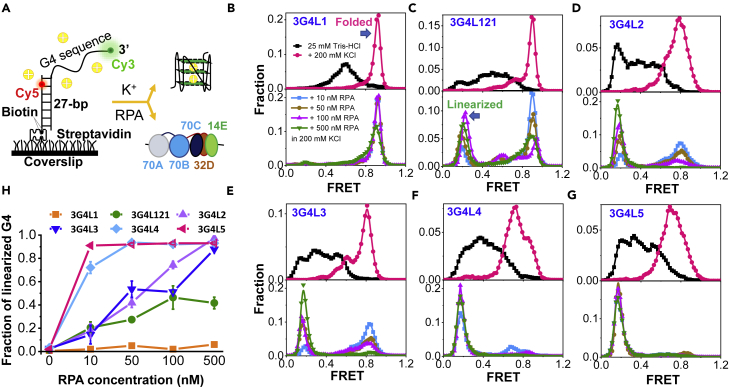
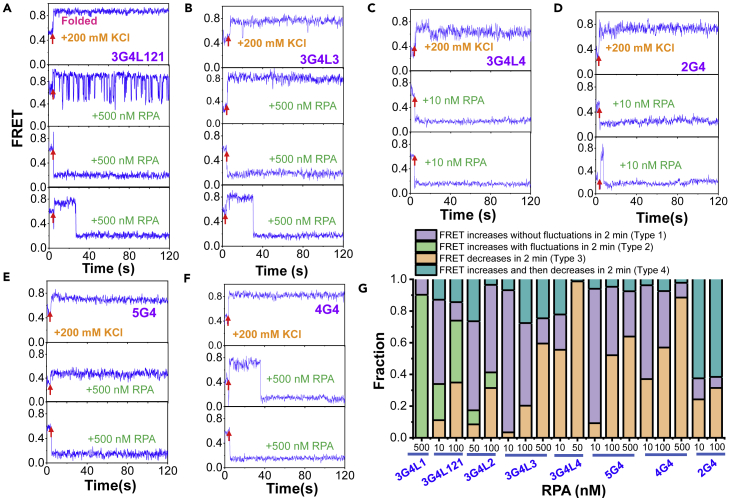
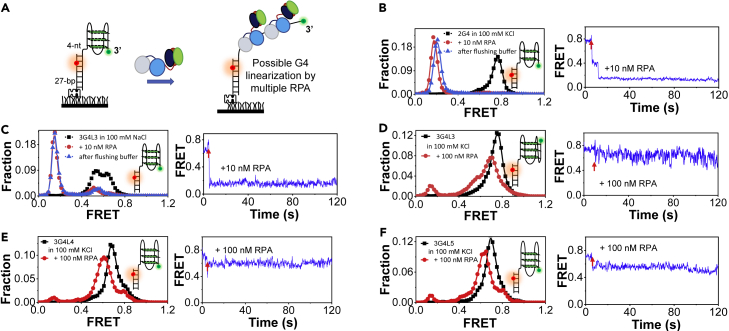
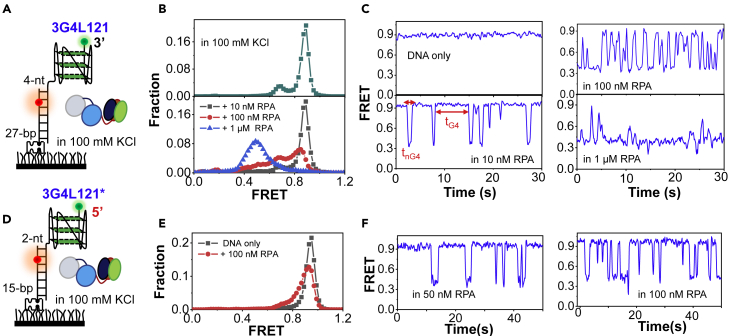
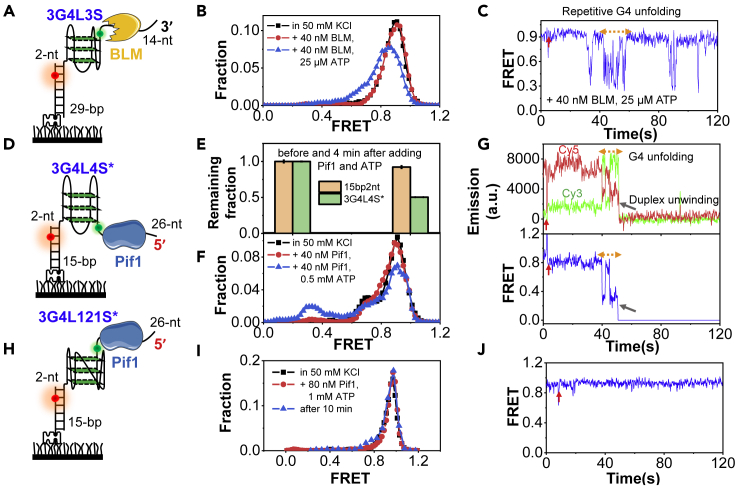
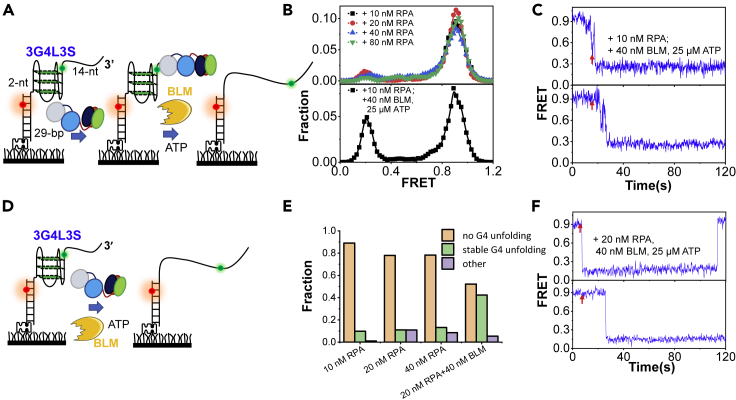
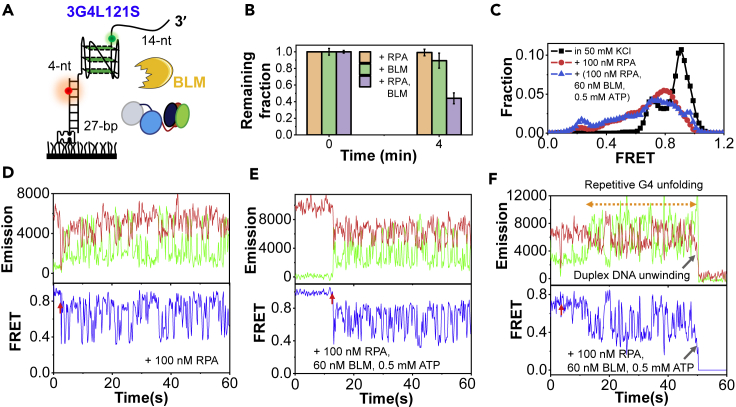
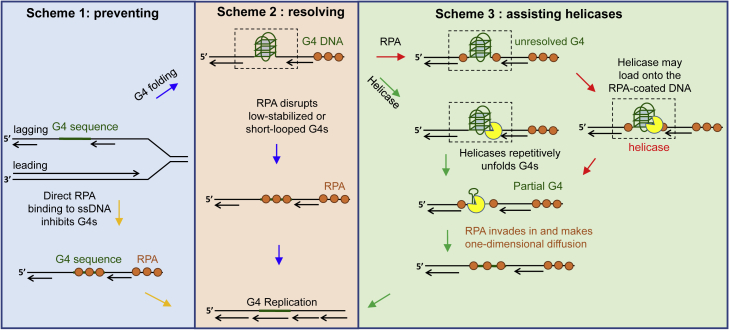
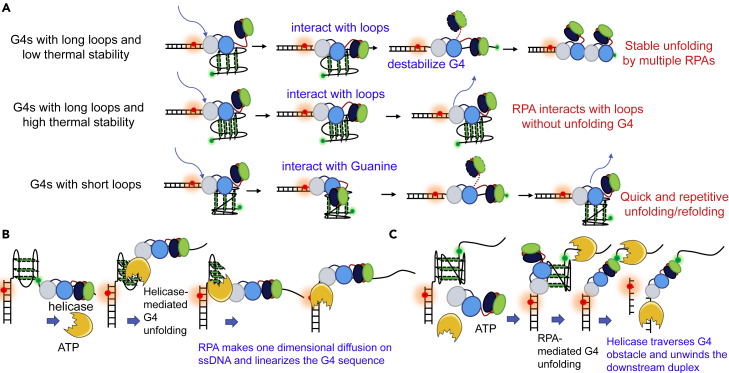
G-quadruplexes (G4s) are non-canonical DNA structures with critical roles in DNA metabolisms. To resolve those structures that can cause replication fork stalling and genomic instability, single-stranded DNA-binding proteins and helicases are required. Here, we characterized the interplay between RPA and helicases on G4s using single-molecule FRET. We first discovered that human RPA efficiently prevents G4 formation by preempting ssDNA before its folding. RPA also differentially interacts with the folded G4s. However, helicases such as human BLM and yeast Pif1 have different G4 preferences from RPA mainly based on loop lengths. More importantly, both RPA and these helicases are required for the stable G4 unfolding, as RPA promotes helicase-mediated repetitive unfolding into durative linear state. Furthermore, BLM can traverse G4 obstacles temporarily disrupted by RPA and continue to unwind downstream duplex. We finally proposed the mechanisms underlying above functions of RPA in preventing, resolving, and assisting helicases to eliminate G4s.
Keywords: Molecular biology; Molecular structure.
© 2021 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
