

Microtubule reorganization during female meiosis in C. elegans
- PMID: 34114562
- PMCID: PMC8225387
- DOI: 10.7554/eLife.58903
Microtubule reorganization during female meiosis in C. elegans
Erratum in
-
Correction: Microtubule reorganization during female meiosis in C. elegans.Elife. 2023 Feb 23;12:e86893. doi: 10.7554/eLife.86893. Elife. 2023. PMID: 36820823 Free PMC article.
Abstract
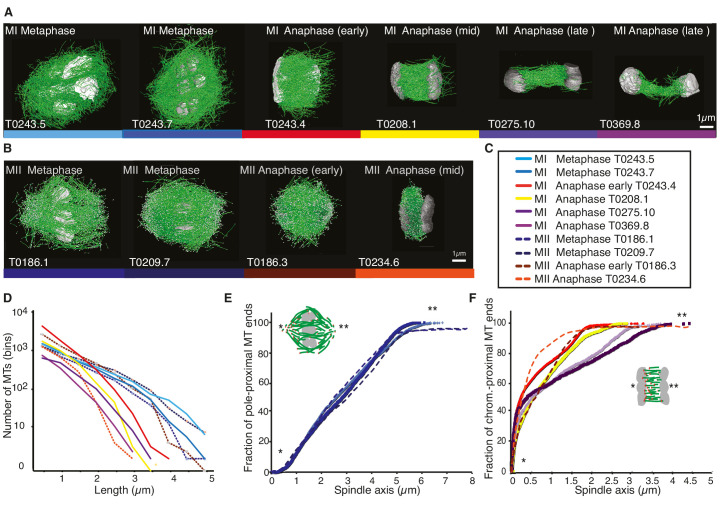
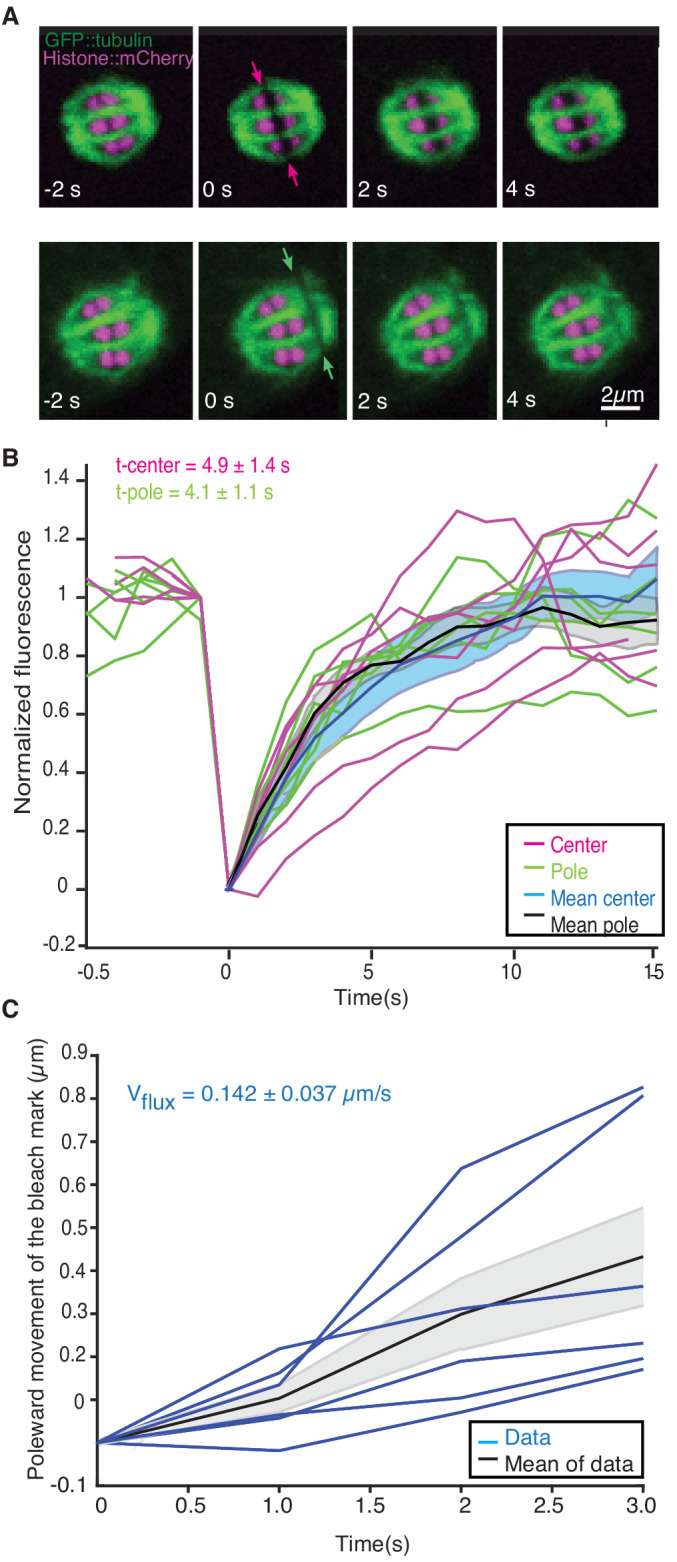
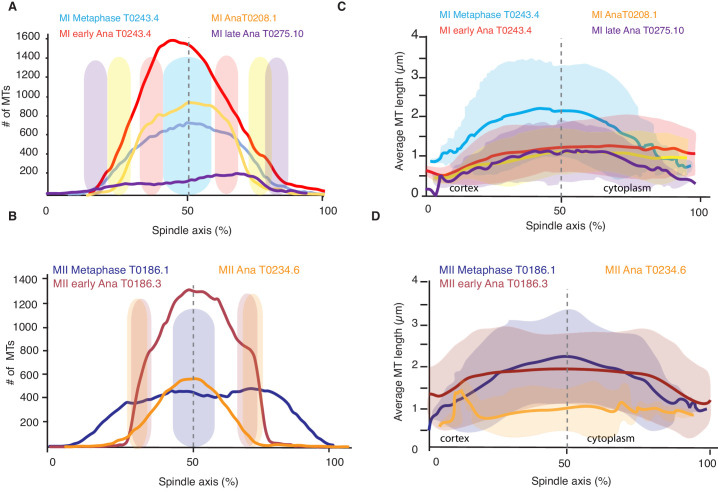
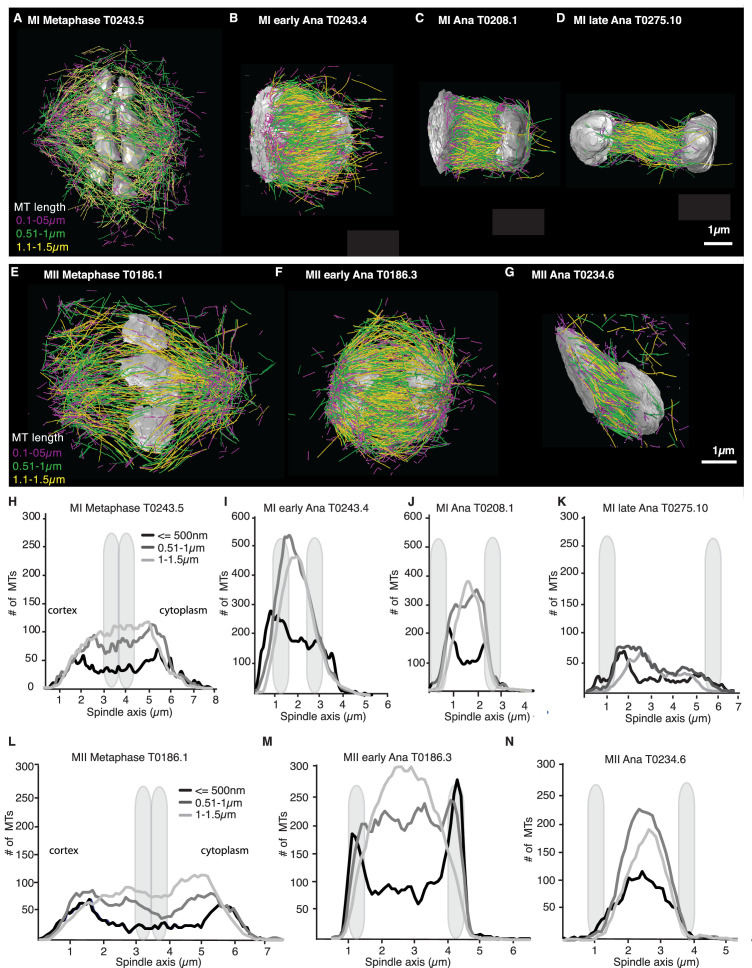
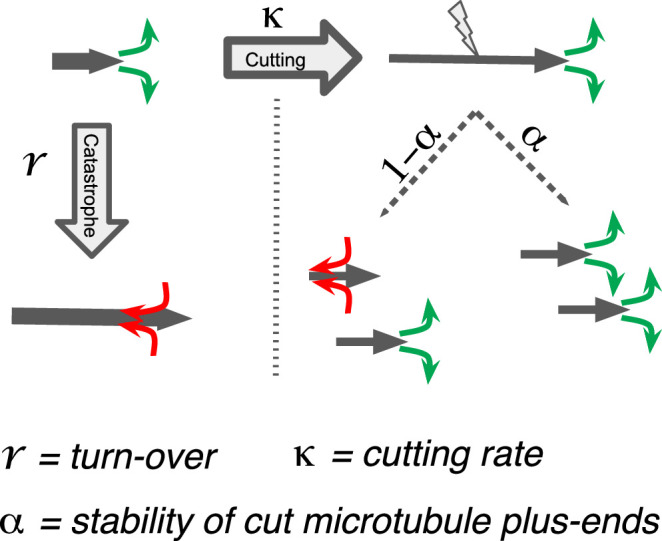
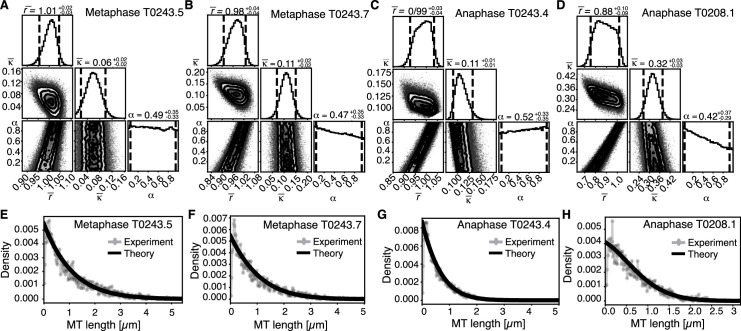
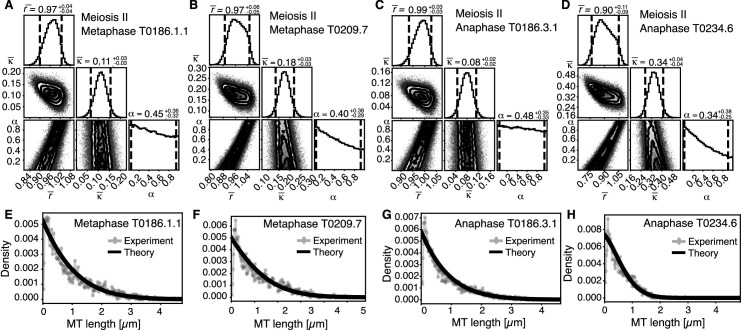
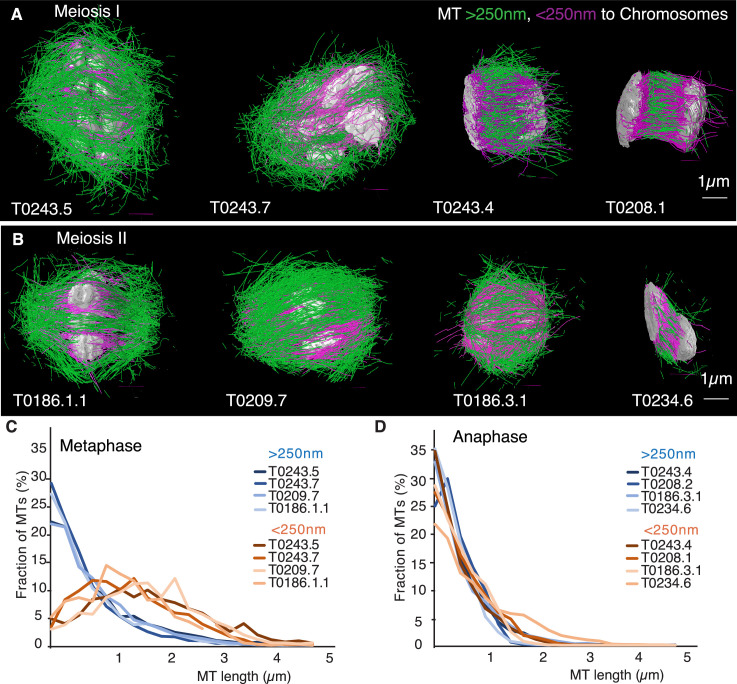
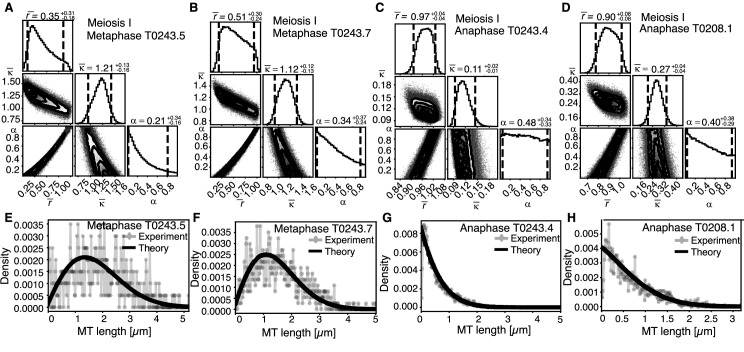
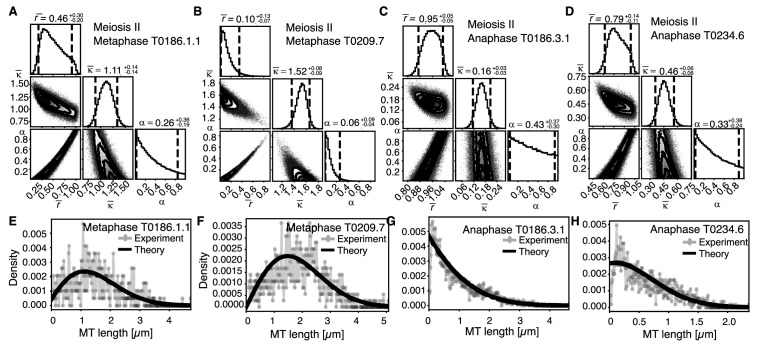
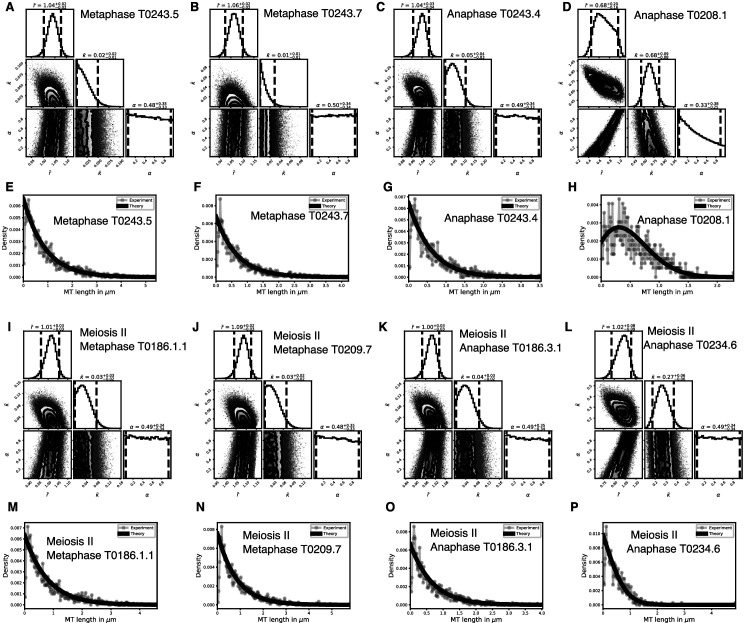
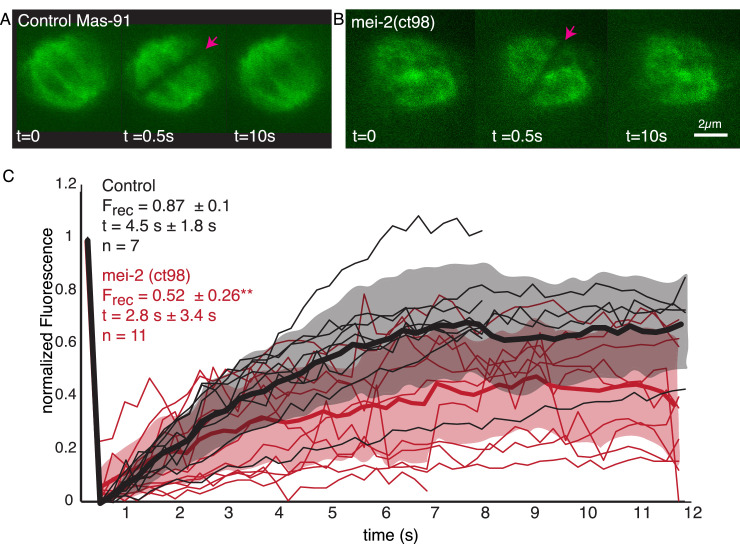
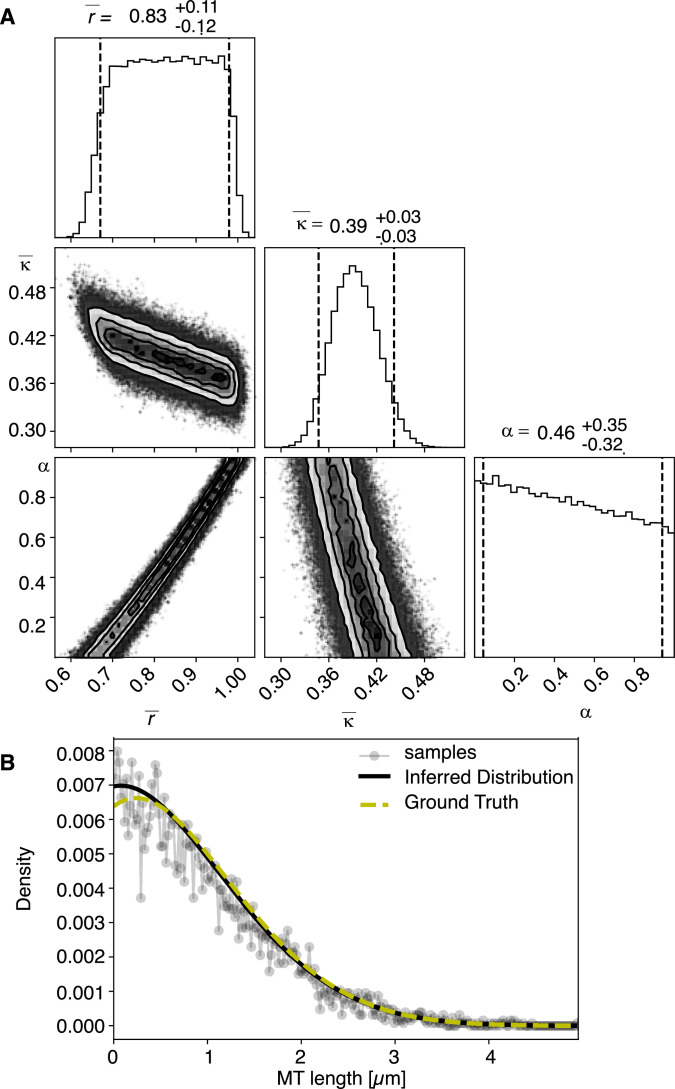
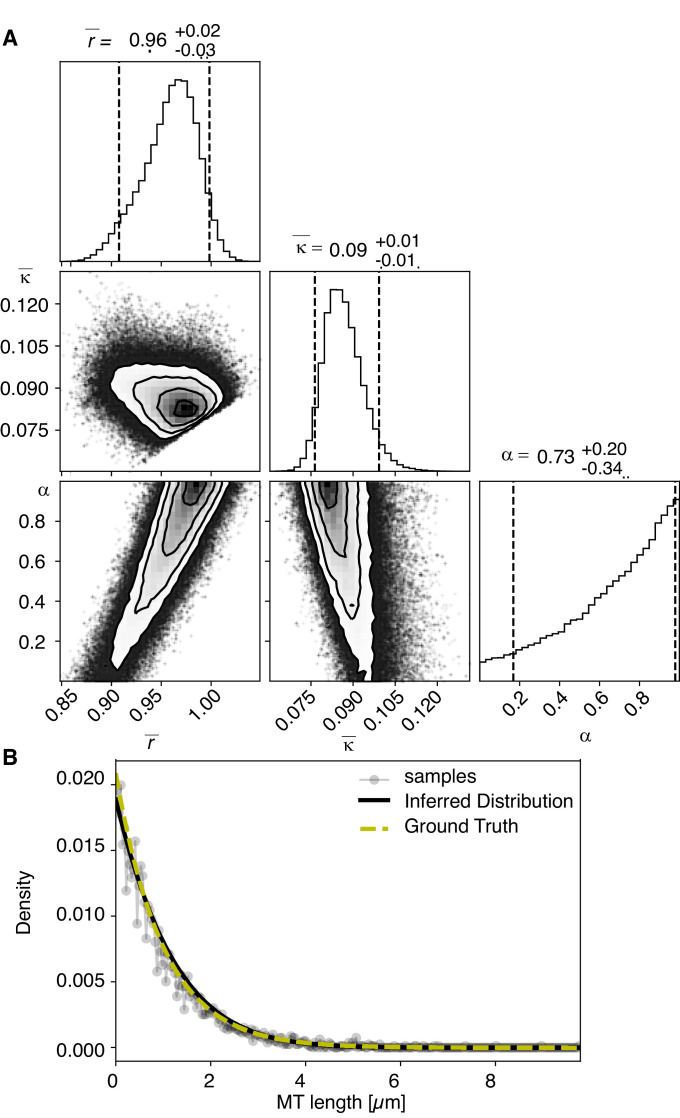
Most female meiotic spindles undergo striking morphological changes while transitioning from metaphase to anaphase. The ultra-structure of meiotic spindles, and how changes to this structure correlate with such dramatic spindle rearrangements remains largely unknown. To address this, we applied light microscopy, large-scale electron tomography and mathematical modeling of female meiotic Caenorhabditis elegans spindles. Combining these approaches, we find that meiotic spindles are dynamic arrays of short microtubules that turn over within seconds. The results show that the metaphase to anaphase transition correlates with an increase in microtubule numbers and a decrease in their average length. Detailed analysis of the tomographic data revealed that the microtubule length changes significantly during the metaphase-to-anaphase transition. This effect is most pronounced for microtubules located within 150 nm of the chromosome surface. To understand the mechanisms that drive this transition, we developed a mathematical model for the microtubule length distribution that considers microtubule growth, catastrophe, and severing. Using Bayesian inference to compare model predictions and data, we find that microtubule turn-over is the major driver of the spindle reorganizations. Our data suggest that in metaphase only a minor fraction of microtubules, those closest to the chromosomes, are severed. The large majority of microtubules, which are not in close contact with chromosomes, do not undergo severing. Instead, their length distribution is fully explained by growth and catastrophe. This suggests that the most prominent drivers of spindle rearrangements are changes in nucleation and catastrophe rate. In addition, we provide evidence that microtubule severing is dependent on katanin.
Keywords: C. elegans; Meiosis; cell biology; computational biology; mathematical modelling; microtubules; systems biology; tomography.
© 2021, Lantzsch et al.
Conflict of interest statement
IL, CY, YC, VZ, HY, NL, ES, AP, SP, MS, SF, SR No competing interests declared
Figures
Similar articles
-
Katanin maintains meiotic metaphase chromosome alignment and spindle structure in vivo and has multiple effects on microtubules in vitro.Mol Biol Cell. 2014 Apr;25(7):1037-49. doi: 10.1091/mbc.E13-12-0764. Epub 2014 Feb 5. Mol Biol Cell. 2014. PMID: 24501424 Free PMC article.
-
MEI-1/katanin is required for translocation of the meiosis I spindle to the oocyte cortex in C elegans.Dev Biol. 2003 Aug 1;260(1):245-59. doi: 10.1016/s0012-1606(03)00216-1. Dev Biol. 2003. PMID: 12885567
-
A Switch in Microtubule Orientation during C. elegans Meiosis.Curr Biol. 2018 Sep 24;28(18):2991-2997.e2. doi: 10.1016/j.cub.2018.07.012. Epub 2018 Sep 6. Curr Biol. 2018. PMID: 30197085
-
Oocyte Meiotic Spindle Assembly and Function.Curr Top Dev Biol. 2016;116:65-98. doi: 10.1016/bs.ctdb.2015.11.031. Epub 2016 Jan 23. Curr Top Dev Biol. 2016. PMID: 26970614 Free PMC article. Review.
-
The chromosomal basis of meiotic acentrosomal spindle assembly and function in oocytes.Chromosoma. 2017 Jun;126(3):351-364. doi: 10.1007/s00412-016-0618-1. Epub 2016 Nov 11. Chromosoma. 2017. PMID: 27837282 Free PMC article. Review.
Cited by
-
An intelligent workflow for sub-nanoscale 3D reconstruction of intact synapses from serial section electron tomography.BMC Biol. 2023 Sep 25;21(1):198. doi: 10.1186/s12915-023-01696-x. BMC Biol. 2023. PMID: 37743470 Free PMC article.
-
Roles of Tubulin Concentration during Prometaphase and Ran-GTP during Anaphase of C. elegans meiosis.bioRxiv [Preprint]. 2024 Jun 25:2024.04.19.590357. doi: 10.1101/2024.04.19.590357. bioRxiv. 2024. Update in: Life Sci Alliance. 2024 Jul 3;7(9):e202402884. doi: 10.26508/lsa.202402884. PMID: 38659754 Free PMC article. Updated. Preprint.
-
Microtubule length correlates with spindle length in C. elegans meiosis.Cytoskeleton (Hoboken). 2024 Aug;81(8):356-368. doi: 10.1002/cm.21849. Epub 2024 Mar 7. Cytoskeleton (Hoboken). 2024. PMID: 38450962 Free PMC article.
-
Redundant microtubule crosslinkers prevent meiotic spindle bending to ensure diploid offspring in C. elegans.PLoS Genet. 2023 Dec 27;19(12):e1011090. doi: 10.1371/journal.pgen.1011090. eCollection 2023 Dec. PLoS Genet. 2023. PMID: 38150489 Free PMC article.
-
Microtubules in Martini: Parameterizing a heterogeneous elastic-network towards a mechanically accurate microtubule.PNAS Nexus. 2025 Jun 21;4(7):pgaf202. doi: 10.1093/pnasnexus/pgaf202. eCollection 2025 Jul. PNAS Nexus. 2025. PMID: 40599826 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
