

Perilla (Perilla frutescens) leaf extract inhibits SARS-CoV-2 via direct virus inactivation
- PMID: 34119448
- PMCID: PMC7840404
- DOI: 10.1016/j.bj.2021.01.005
Perilla (Perilla frutescens) leaf extract inhibits SARS-CoV-2 via direct virus inactivation
Abstract
Background: While severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection presents with mild or no symptoms in most cases, a significant number of patients become critically ill. Remdesivir has been approved for the treatment of coronavirus disease 2019 (COVID-19) in several countries, but its use as monotherapy has not substantially lowered mortality rates. Because agents from traditional Chinese medicine (TCM) have been successfully utilized to treat pandemic and endemic diseases, we designed the current study to identify novel anti-SARS-CoV-2 agents from TCM.
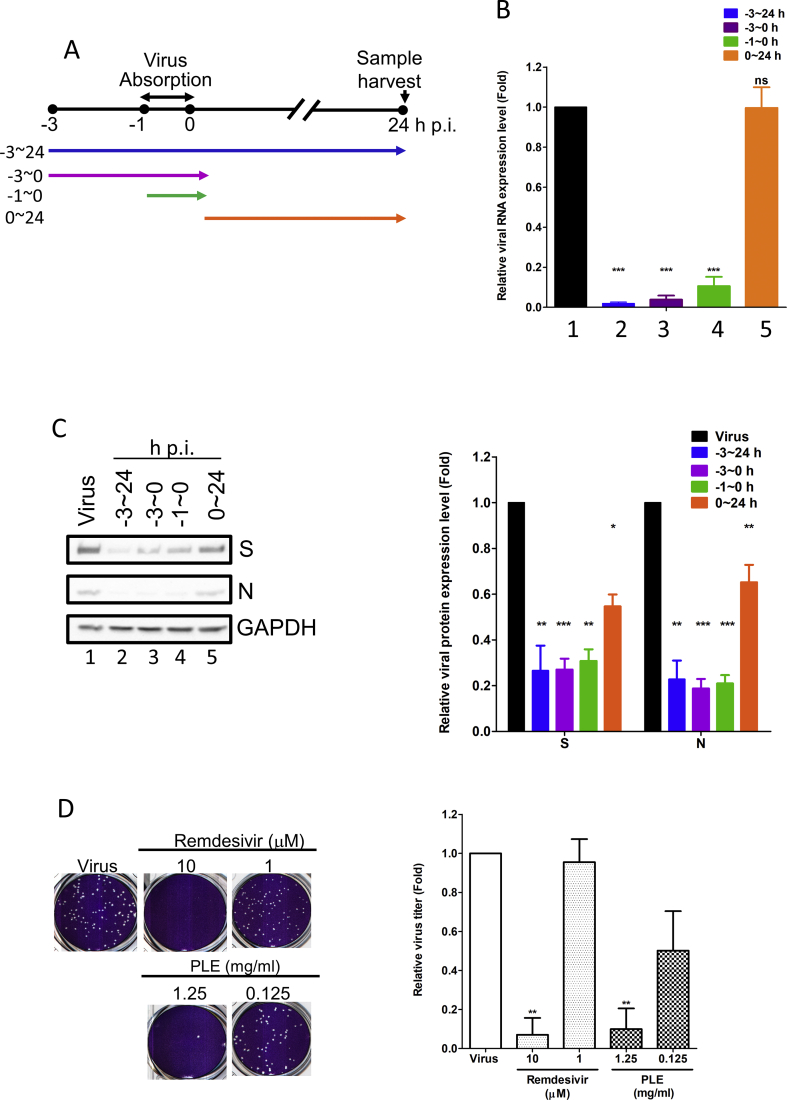
Methods: We initially used an antivirus-induced cell death assay to screen a panel of herbal extracts. The inhibition of the viral infection step was investigated through a time-of-drug-addition assay, whereas a plaque reduction assay was carried out to validate the antiviral activity. Direct interaction of the candidate TCM compound with viral particles was assessed using a viral inactivation assay. Finally, the potential synergistic efficacy of remdesivir and the TCM compound was examined with a combination assay.
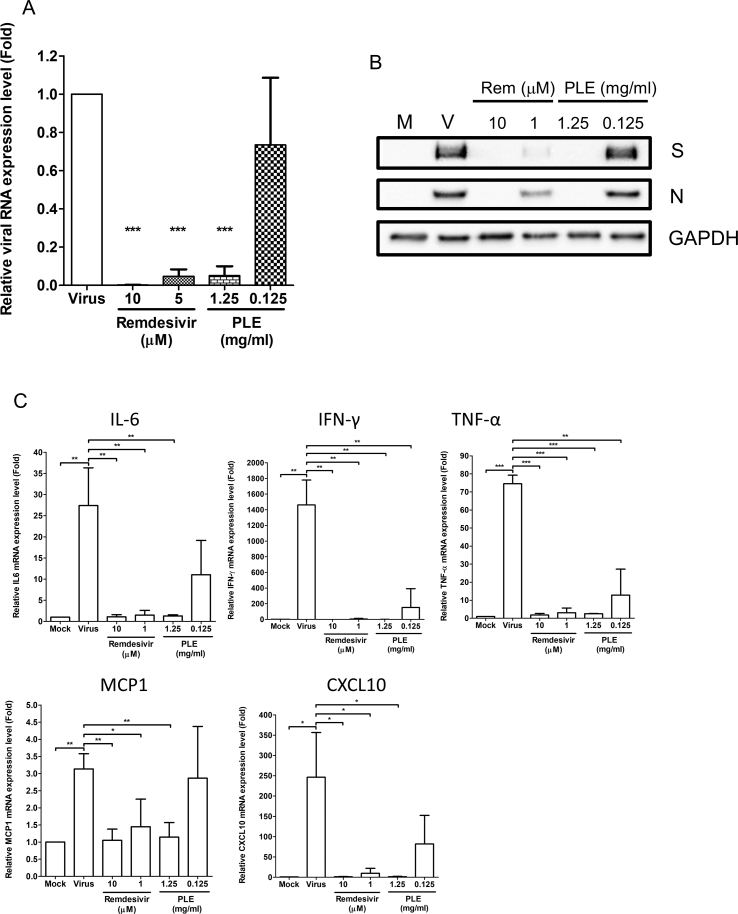
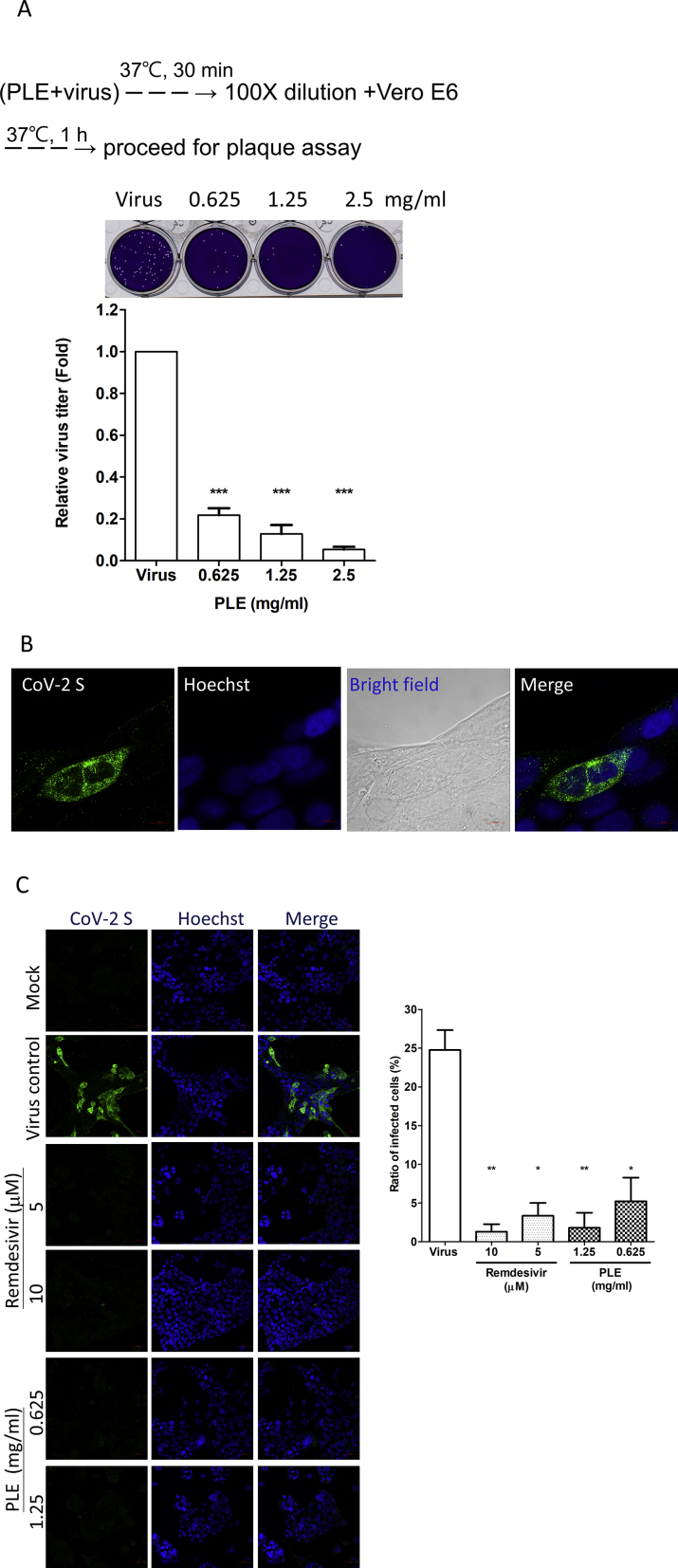
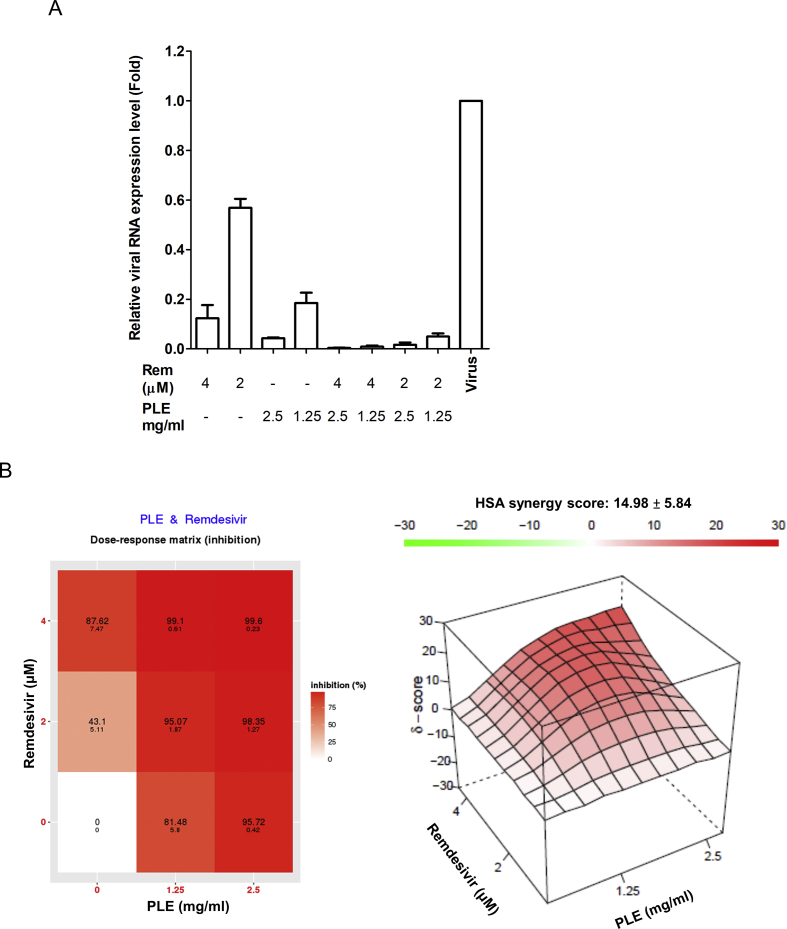
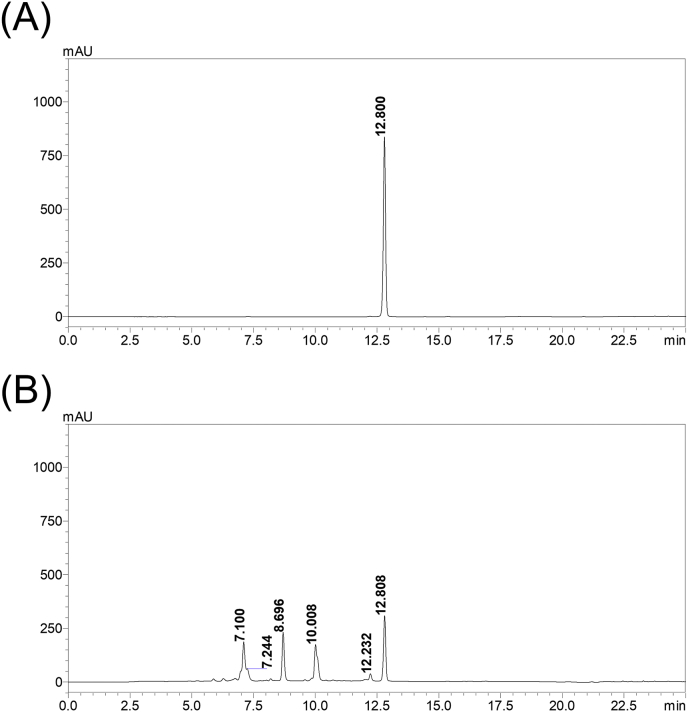
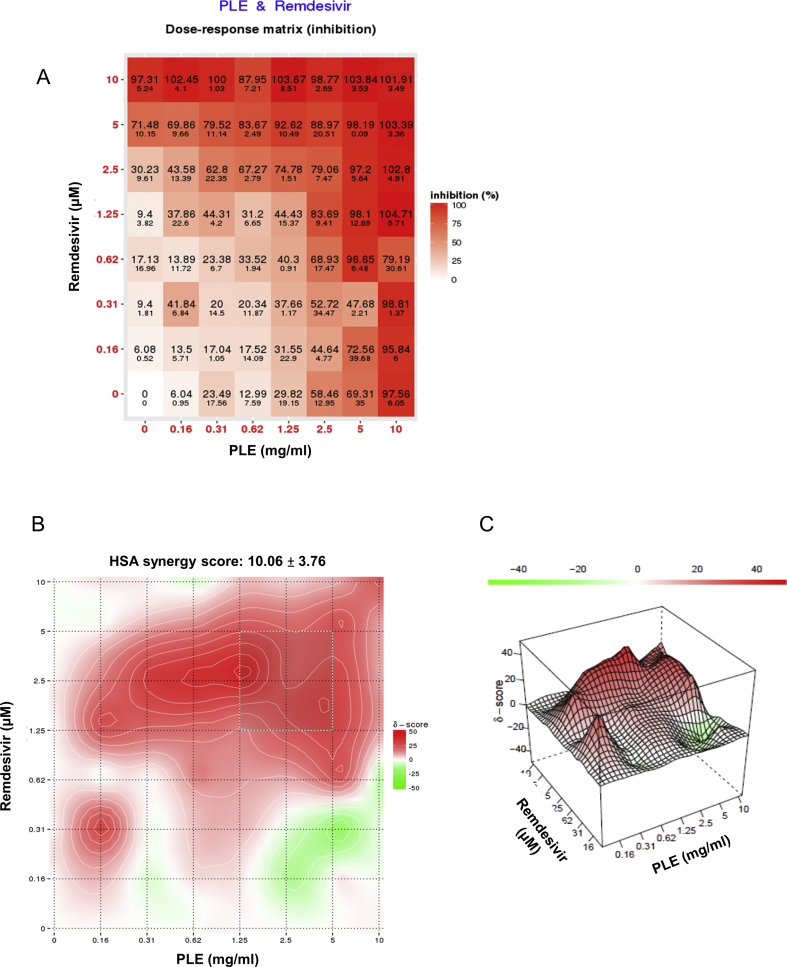
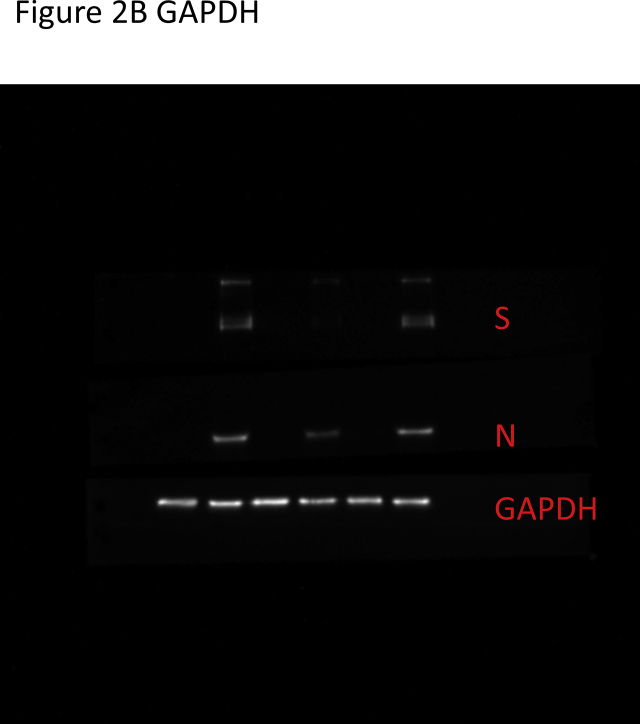
Results: The herbal medicine Perilla leaf extract (PLE, approval number 022427 issued by the Ministry of Health and Welfare, Taiwan) had EC50 of 0.12 ± 0.06 mg/mL against SARS-CoV-2 in Vero E6 cells - with a selectivity index of 40.65. Non-cytotoxic PLE concentrations were capable of blocking viral RNA and protein synthesis. In addition, they significantly decreased virus-induced cytokine release and viral protein/RNA levels in the human lung epithelial cell line Calu-3. PLE inhibited viral replication by inactivating the virion and showed additive-to-synergistic efficacy against SARS-CoV-2 when used in combination with remdesivir.
Conclusion: Our results demonstrate for the first time that PLE is capable of inhibiting SARS-CoV-2 replication by inactivating the virion. Our data may prompt additional investigation on the clinical usefulness of PLE for preventing or treating COVID-19.
Keywords: COVID-19; Coronavirus; Perilla frutescens (L.) Britt; SARS-CoV-2; Traditional Chinese medicine; Zisu.
Copyright © 2021 Chang Gung University. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare they have no actual or potential competing financial interests.
Figures


References
-
- Masters P.S., Perlman S. Coronaviridae. In: David M., Knipe P.M.H., Martin Malcolm A., Griffin Diane E., Lamb Robert A., Bernard Roizman, Straus Stephen E., editors. Fields virology. Lippincott Williams & Wilkins; Philadelphia: 2013. pp. 825–858.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
