

Molecular characterization and expression analysis of ribosomal L18/L5e gene in Pennisetum glaucum (L.) R. Br
- PMID: 34121902
- PMCID: PMC8176002
- DOI: 10.1016/j.sjbs.2021.03.035
Molecular characterization and expression analysis of ribosomal L18/L5e gene in Pennisetum glaucum (L.) R. Br
Abstract
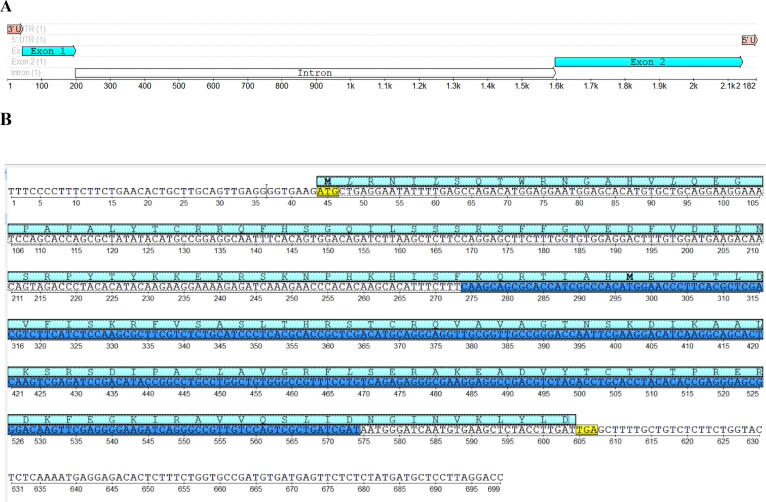
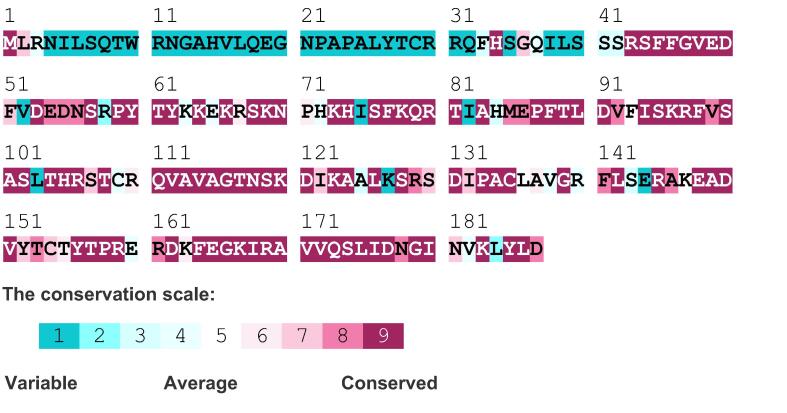
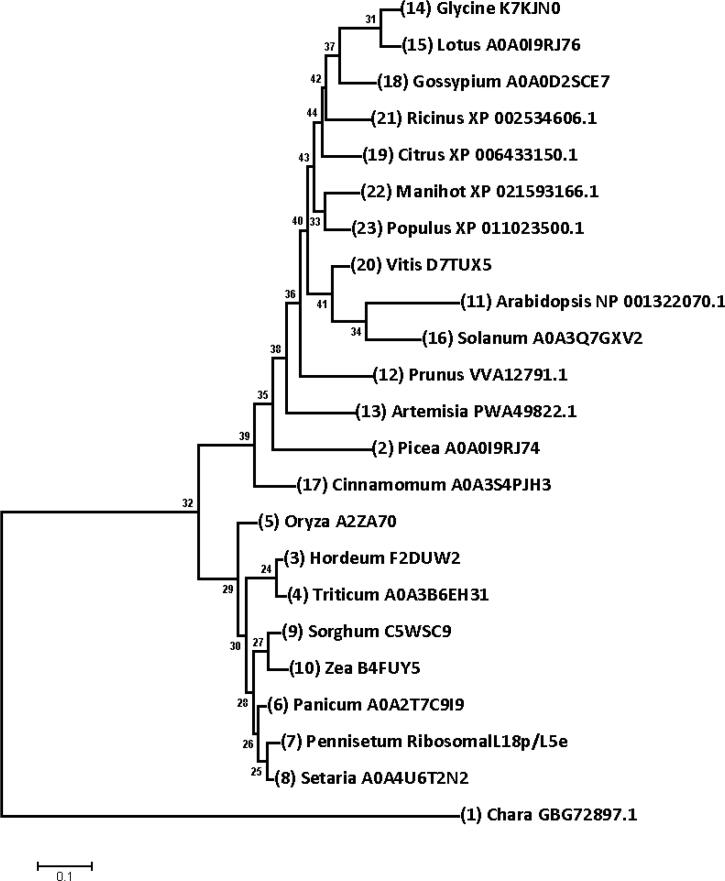
Ribosomal L18/L5e (RL18/L5e) is a member of the ribosomal L18/L5e protein family, which has an essential function in translation of mRNA into protein in the large ribosomal subunit. In this study, RL18/L5e was isolated and sequenced from local Pennisetum glaucum (L.) R. Br. cultivar which is known to adapt to environmental stress. The obtained cDNA for PgRL18/L5e was 699 bp in length, with an open reading frame of 564 bp. The deduced protein sequence contained 187 amino acids and comprised an RL18/L5e domain, which shared high sequence identity with orthologous proteins from Viridiplantae. The obtained PgRL18/L5e cDNA contained two exons of 154 and 545 bp, respectively, and an intron of 1398 bp. Secondary and 3D structures of the deduced PgRL18/L5e protein were predicted using in silico tools. Phylogenetic analysis showed close relationships between the PgRL18/L5e protein and its orthologs from monocot species. Multiple sequence alignment showed high identity in the RL18/L5e domain sequence in all orthologous proteins in Viridiplantae. Moreover, all orthologous RL18/L5e proteins shared the same domain architecture and were nearly equal in length. Quantitative real-time PCR indicated a higher transcript abundance of PgRL18/L5e in shoots than in roots of 3-day-old seedlings. Moreover, the expression of PgRL18/L5e in seedlings under cold and drought stress was substantially lower than that in untreated seedlings, whereas the highest expression was shown under heat stress. This study provides insights into the structure and function of the RL18/L5e gene in tolerant crops, which could facilitate the understanding of the role of the various plant ribosomal proteins in adaptation to extreme environments.
Keywords: Gene expression; Pennisetum glaucum; Phylogenetics; Ribosomal L18/L5e; cDNA.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
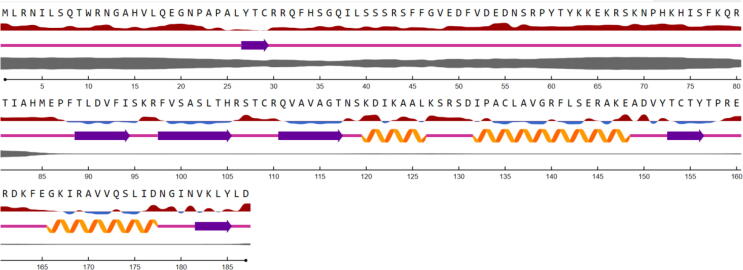
 ; the sequences denoted in red are exposed, and those denoted in blue are buried (threshold at 25%). Secondary structure is illustrated as follows:
; the sequences denoted in red are exposed, and those denoted in blue are buried (threshold at 25%). Secondary structure is illustrated as follows:  Helix,
Helix,  Strand,
Strand,  Coil. The thickness
Coil. The thickness  of the line denotes the probability of disordered residues.
of the line denotes the probability of disordered residues.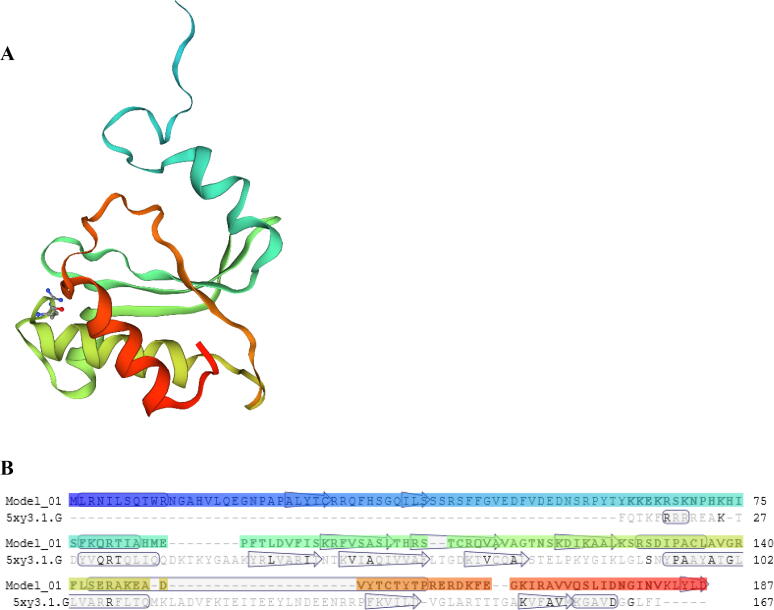

References
-
- Adams K.L., Palmer J.D. Evolution of mitochondrial gene content: gene loss and transfer to the nucleus. Mol. Phylogenet. Evol. 2003;29:380–395. https://doi.org/https://doi.org/10.1016/S1055-7903(03)00194-5. - PubMed
-
- Agarwala R., Barrett T., Beck J., Benson D.A., Bollin C., Bolton E., Bourexis D., Brister J.R., Bryant S.H., Canese K., Cavanaugh M., Charowhas C., Clark K., Dondoshansky I., Feolo M., Fitzpatrick L., Funk K., Geer L.Y., Gorelenkov V., Graeff A., Hlavina W., Holmes B., Johnson M., Kattman B., Khotomlianski V., Kimchi A., Kimelman M., Kimura M., Kitts P., Klimke W., Kotliarov A., Krasnov S., Kuznetsov A., Landrum M.J., Landsman D., Lathrop S., Lee J.M., Leubsdorf C., Lu Z., Madden T.L., Marchler-Bauer A., Malheiro A., Meric P., Karsch-Mizrachi I., Mnev A., Murphy T., Orris R., Ostell J., O’Sullivan C., Palanigobu V., Panchenko A.R., Phan L., Pierov B., Pruitt K.D., Rodarmer K., Sayers E.W., Schneider V., Schoch C.L., Schuler G.D., Sherry S.T., Siyan K., Soboleva A., Soussov V., Starchenko G., Tatusova T.A., Thibaud-Nissen F., Todorov K., Trawick B.W., Vakatov D., Ward M., Yaschenko E., Zasypkin A., Zbicz K. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018;46:D8–D13. doi: 10.1093/nar/gkx1095. - DOI - PMC - PubMed
-
- Ascencio-Ibáñez J.T., Sozzani R., Lee T.J., Chu T.M., Wolfinger R.D., Cella R., Hanley-Bowdoin L. Global analysis of Arabidopsis gene expression uncovers a complex array of changes impacting pathogen response and cell cycle during geminivirus infection. Plant Physiol. 2008;148:436–454. doi: 10.1104/pp.108.121038. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
