

Conservative production of galactosaminogalactan in Metarhizium is responsible for appressorium mucilage production and topical infection of insect hosts
- PMID: 34125872
- PMCID: PMC8224951
- DOI: 10.1371/journal.ppat.1009656
Conservative production of galactosaminogalactan in Metarhizium is responsible for appressorium mucilage production and topical infection of insect hosts
Abstract
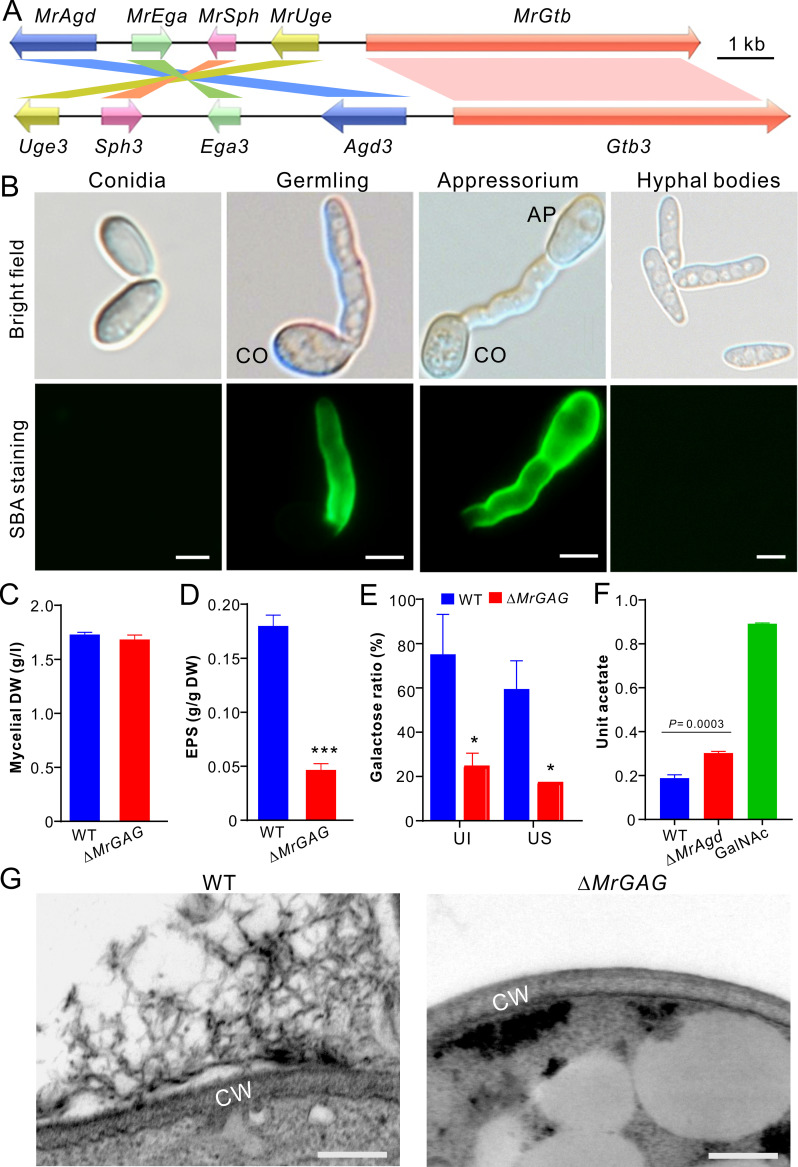
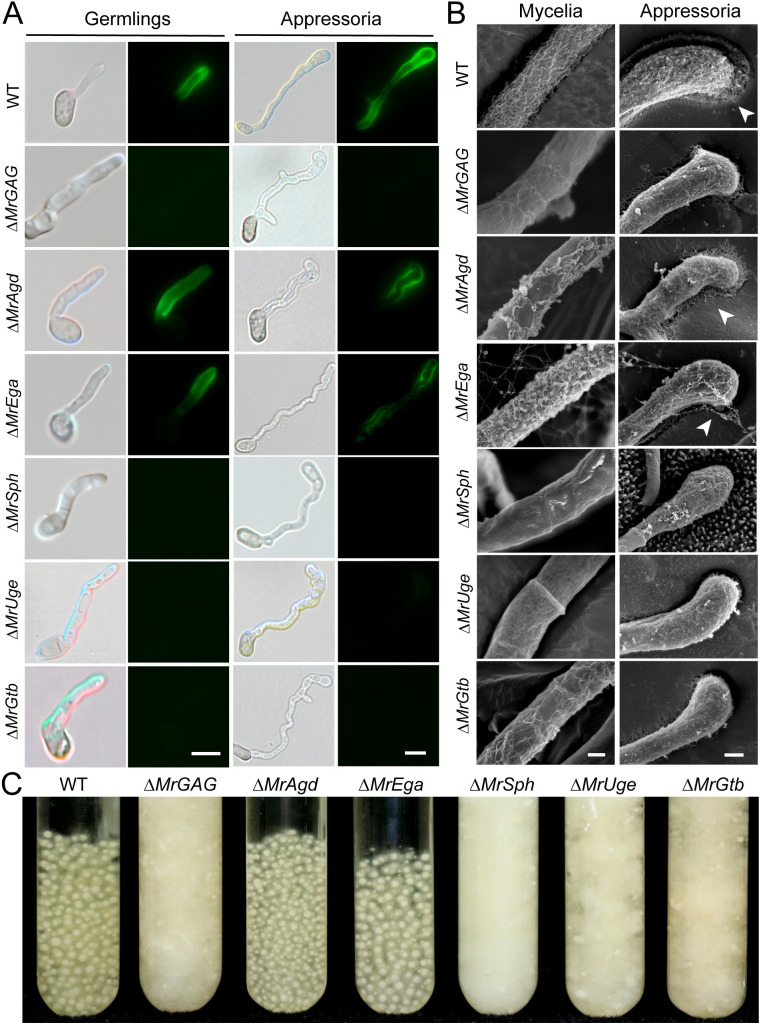
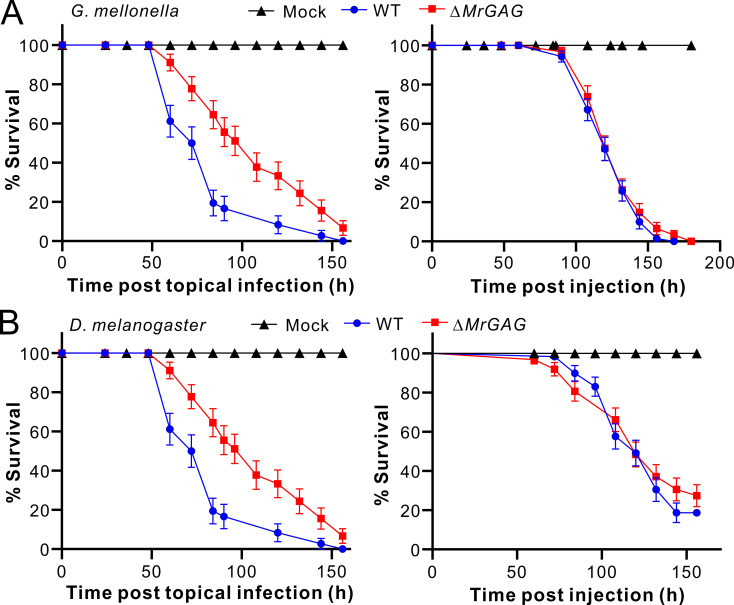
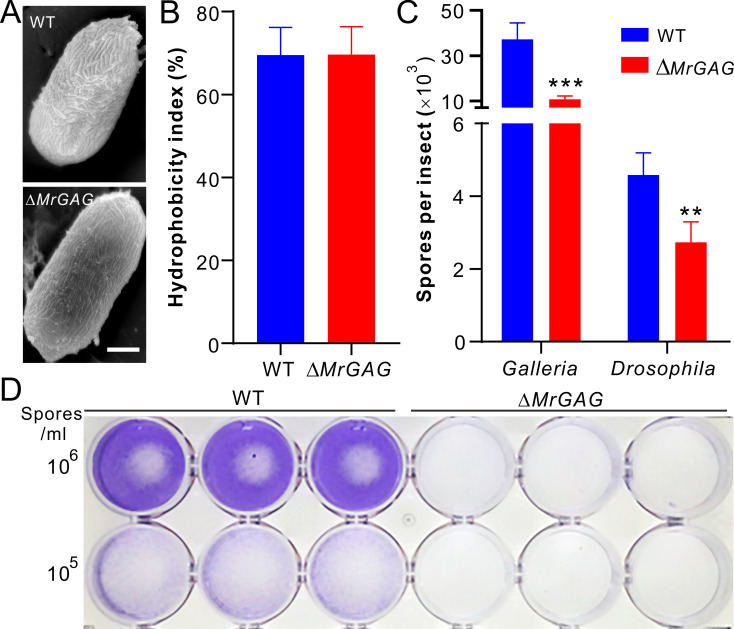
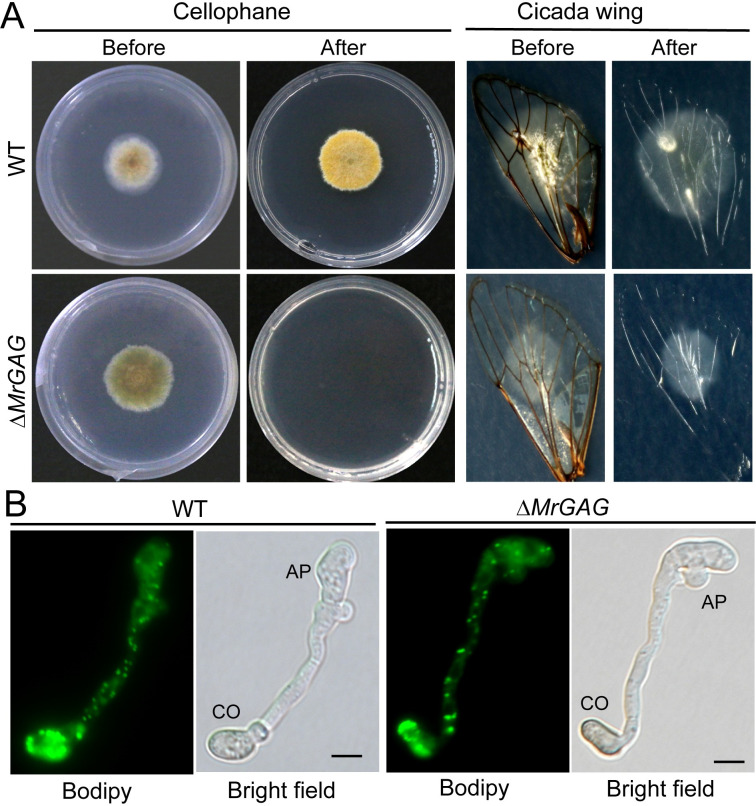
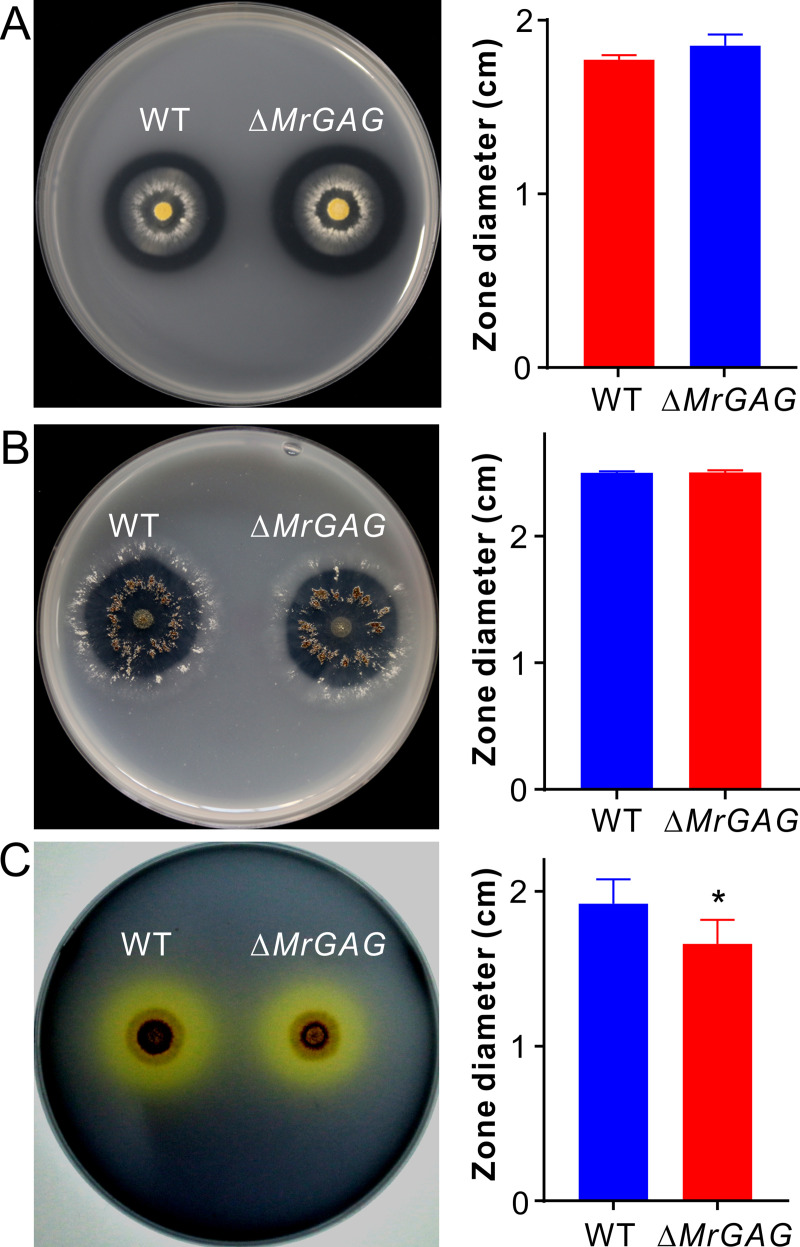
The exopolysaccharide galactosaminogalactan (GAG) has been well characterized in Aspergilli, especially the human pathogen Aspergillus fumigatus. It has been found that a five-gene cluster is responsible for GAG biosynthesis in Aspergilli to mediate fungal adherence, biofilm formation, immunosuppression or induction of host immune defences. Herein, we report the presence of the conserved GAG biosynthetic gene cluster in the insect pathogenic fungus Metarhizium robertsii to mediate either similar or unique biological functions. Deletion of the gene cluster disabled fungal ability to produce GAG on germ tubes, mycelia and appressoria. Relative to the wild type strain, null mutant was impaired in topical infection but not injection of insect hosts. We found that GAG production by Metarhizium is partially acetylated and could mediate fungal adherence to hydrophobic insect cuticles, biofilm formation, and penetration of insect cuticles. In particular, it was first confirmed that this exopolymer is responsible for the formation of appressorium mucilage, the essential extracellular matrix formed along with the infection structure differentiation to mediate cell attachment and expression of cuticle degrading enzymes. In contrast to its production during A. fumigatus invasive growth, GAG is not produced on the Metarhizium cells harvested from insect hemocoels; however, the polymer can glue germ tubes into aggregates to form mycelium pellets in liquid culture. The results of this study unravel the biosynthesis and unique function of GAG in a fungal system apart from the aspergilli species.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
