

IL-13 deficiency exacerbates lung damage and impairs epithelial-derived type 2 molecules during nematode infection
- PMID: 34127548
- PMCID: PMC8321663
- DOI: 10.26508/lsa.202001000
IL-13 deficiency exacerbates lung damage and impairs epithelial-derived type 2 molecules during nematode infection
Abstract
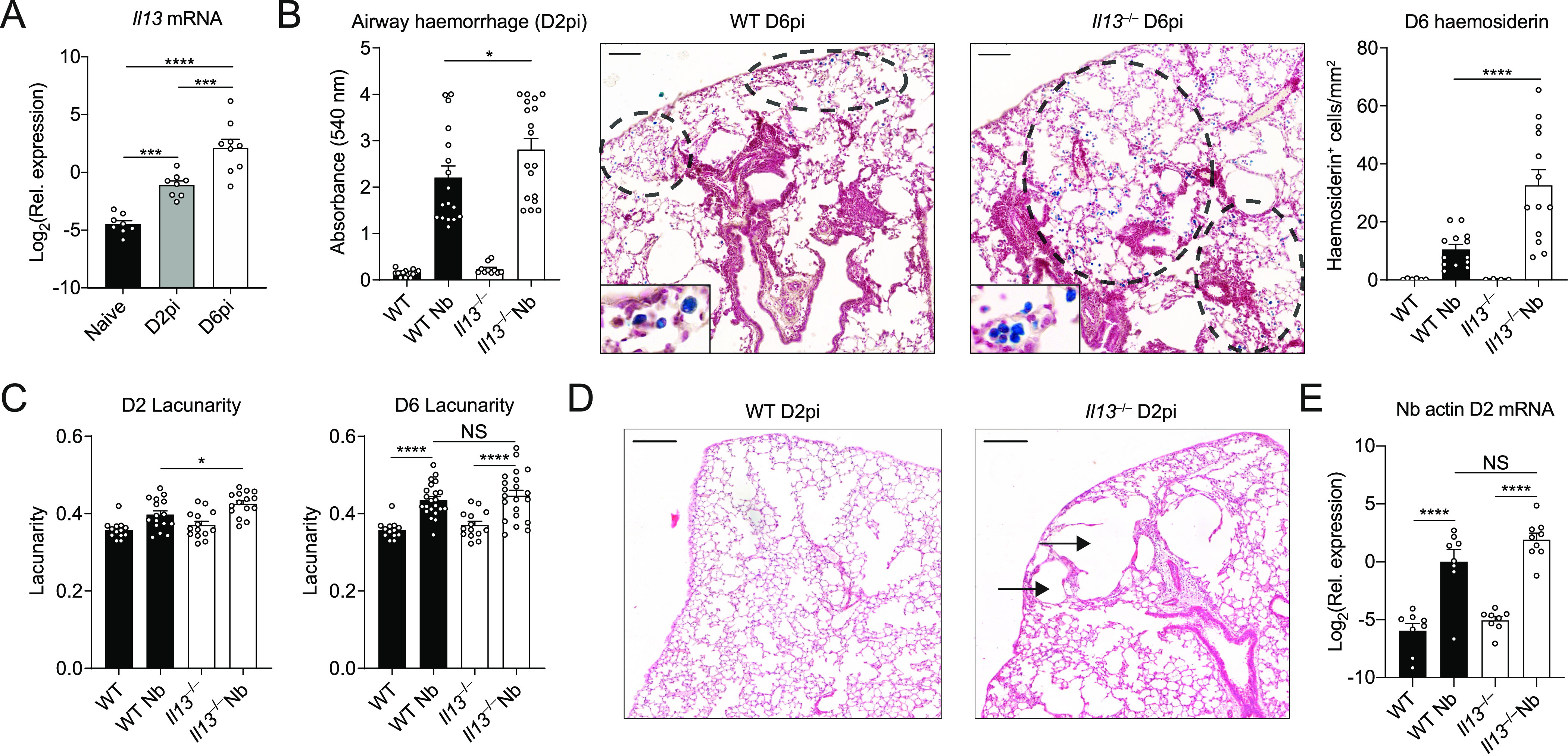
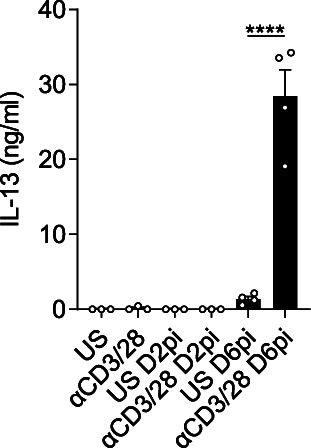
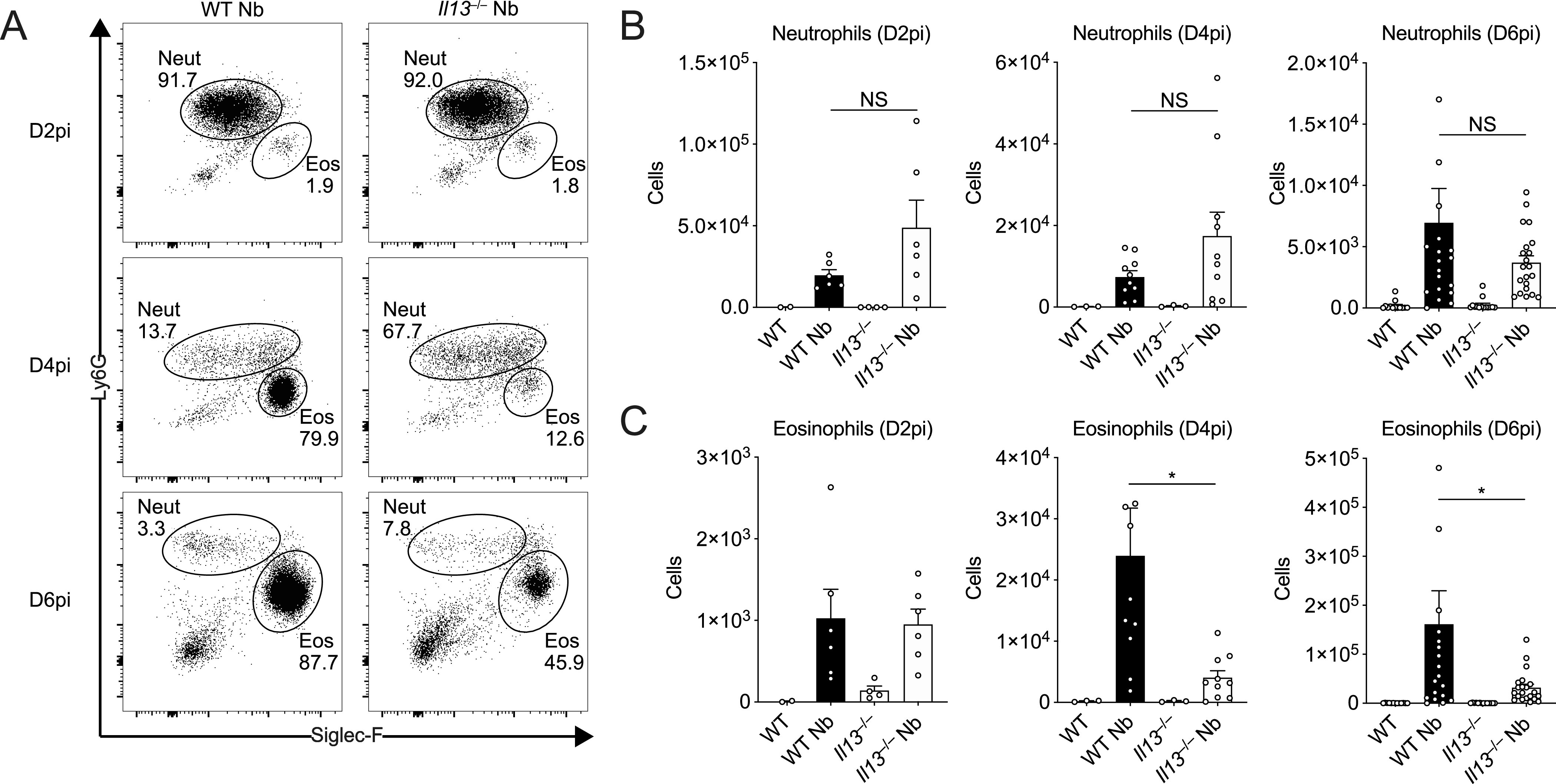
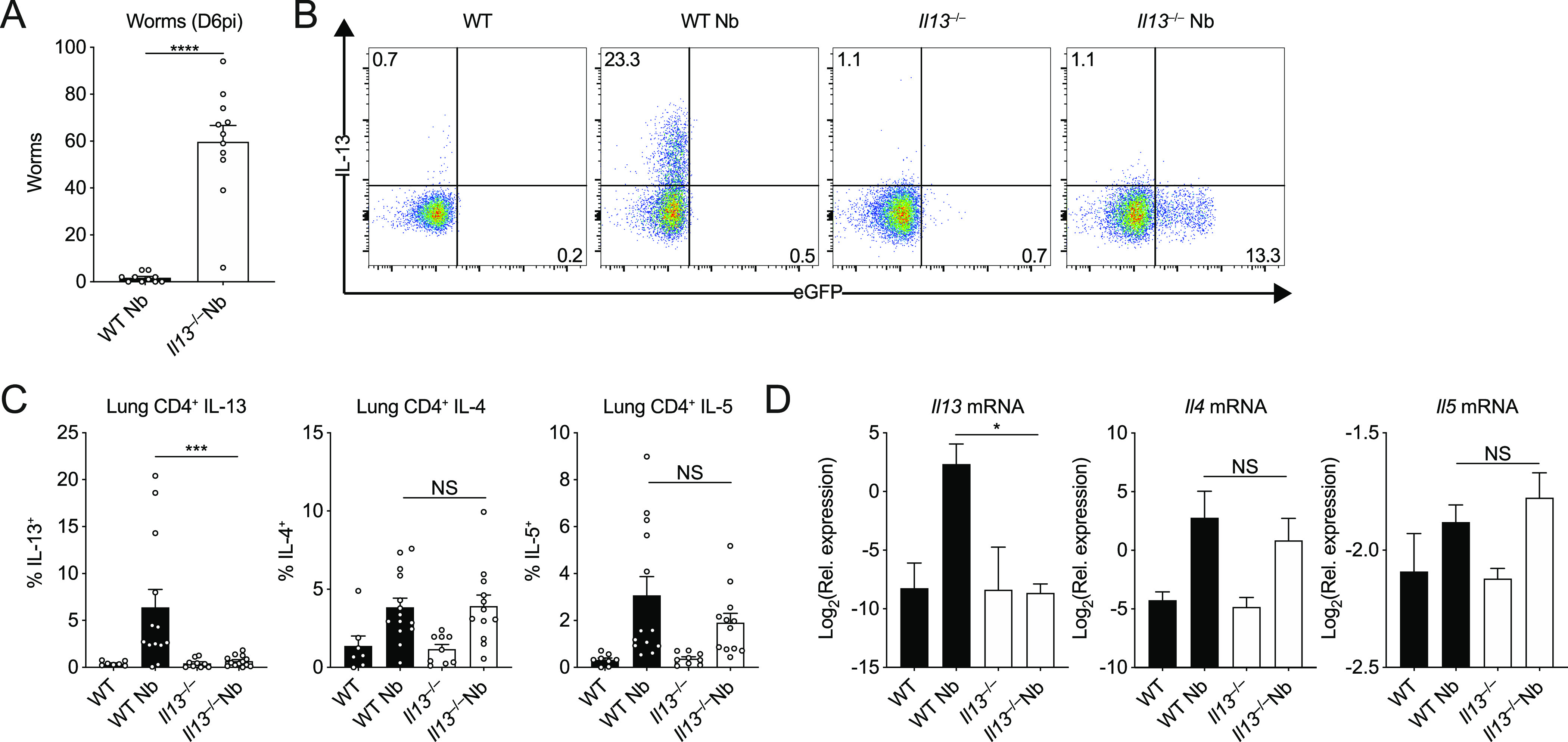
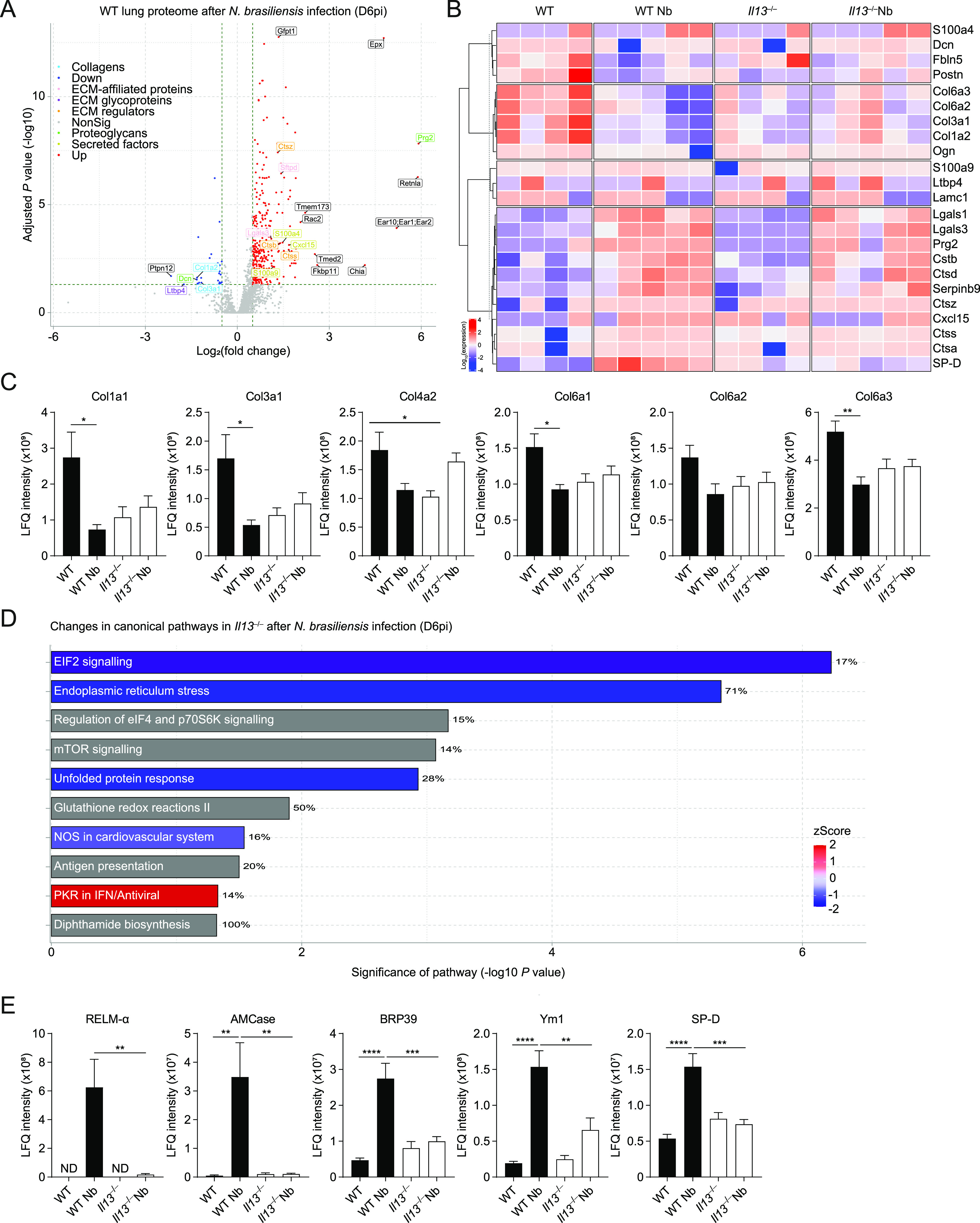
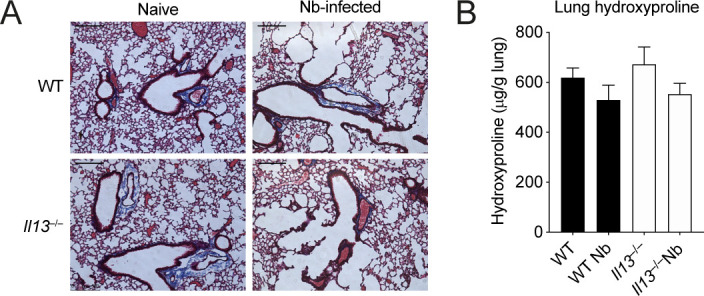
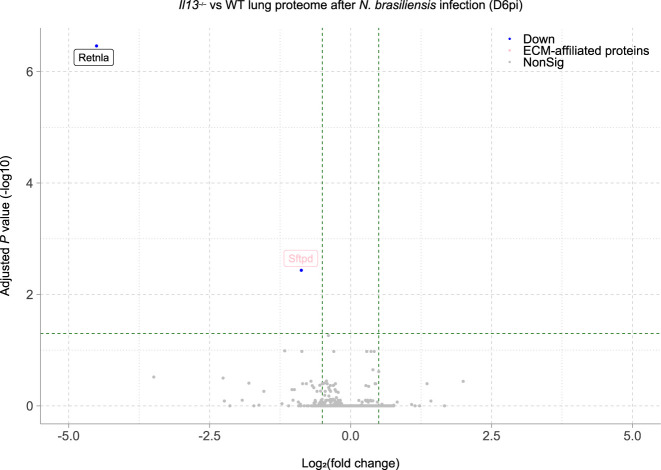
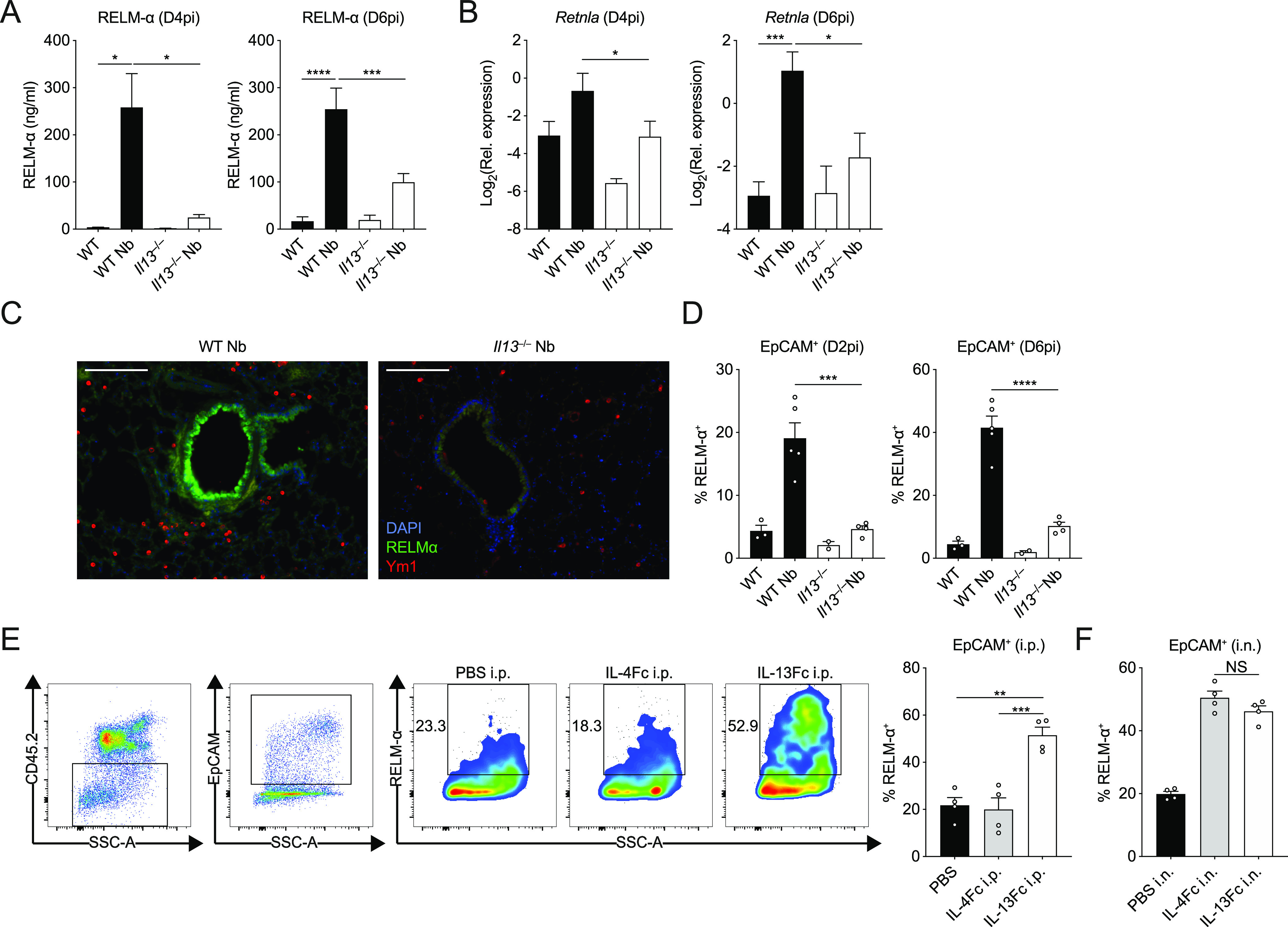
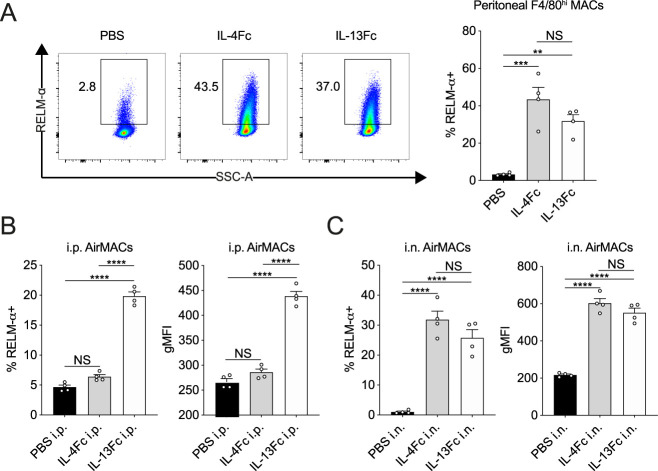
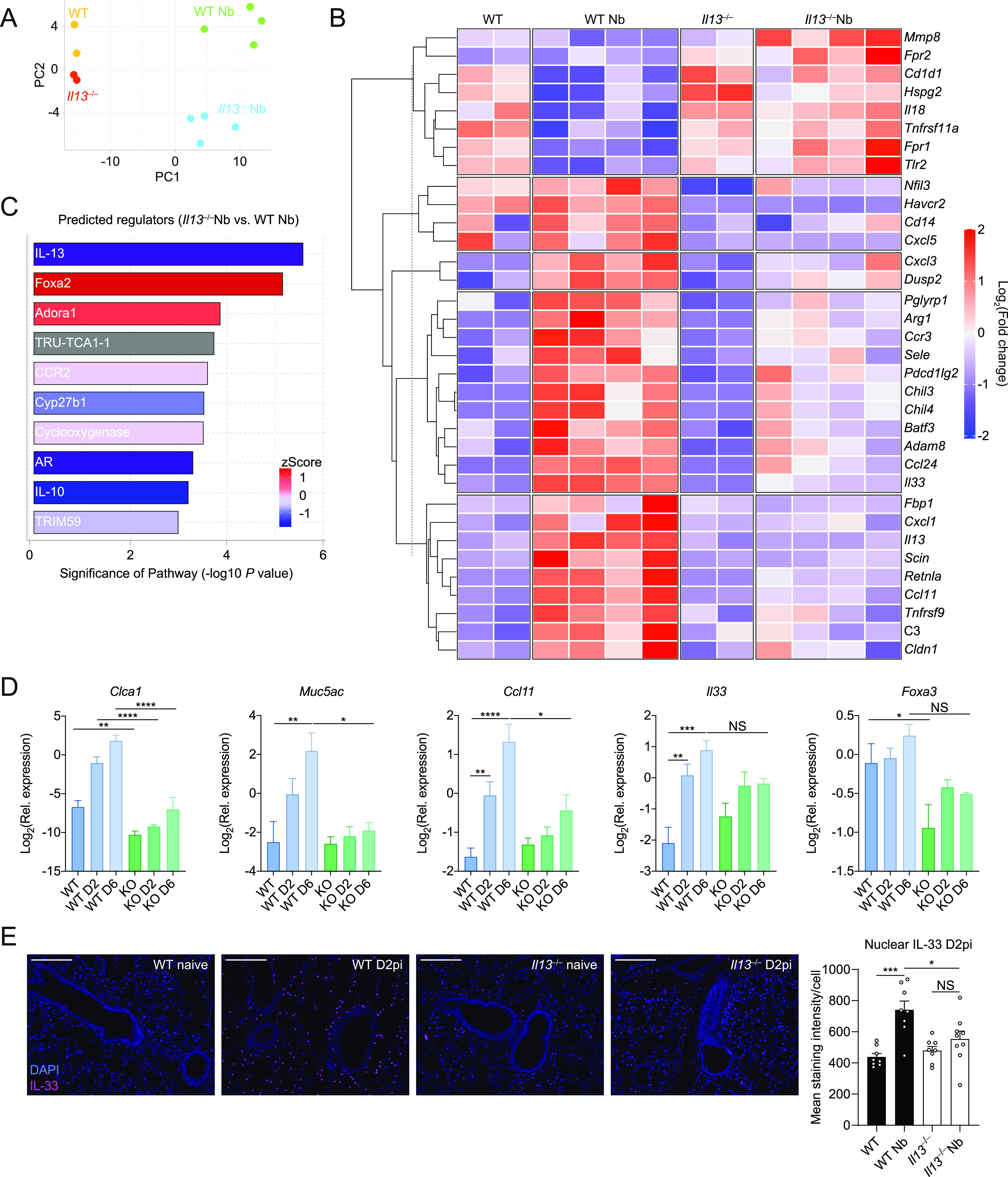
IL-13 is implicated in effective repair after acute lung injury and the pathogenesis of chronic diseases such as allergic asthma. Both these processes involve matrix remodelling, but understanding the specific contribution of IL-13 has been challenging because IL-13 shares receptors and signalling pathways with IL-4. Here, we used Nippostrongylus brasiliensis infection as a model of acute lung damage comparing responses between WT and IL-13-deficient mice, in which IL-4 signalling is intact. We found that IL-13 played a critical role in limiting tissue injury and haemorrhaging in the lung, and through proteomic and transcriptomic profiling, identified IL-13-dependent changes in matrix and associated regulators. We further showed a requirement for IL-13 in the induction of epithelial-derived type 2 effector molecules such as RELM-α and surfactant protein D. Pathway analyses predicted that IL-13 induced cellular stress responses and regulated lung epithelial cell differentiation by suppression of Foxa2 pathways. Thus, in the context of acute lung damage, IL-13 has tissue-protective functions and regulates epithelial cell responses during type 2 immunity.
© 2021 Chenery et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases