

Higher-order organization of biomolecular condensates
- PMID: 34129784
- PMCID: PMC8205532
- DOI: 10.1098/rsob.210137
Higher-order organization of biomolecular condensates
Abstract
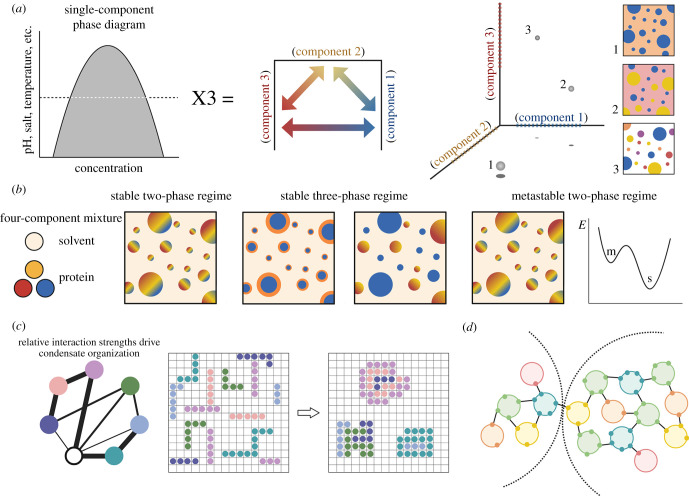
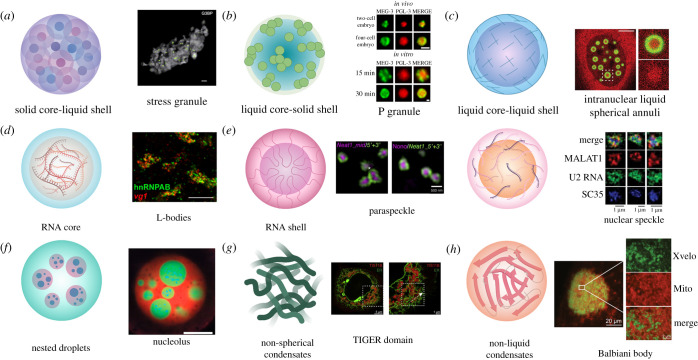
A guiding principle of biology is that biochemical reactions must be organized in space and time. One way this spatio-temporal organization is achieved is through liquid-liquid phase separation (LLPS), which generates biomolecular condensates. These condensates are dynamic and reactive, and often contain a complex mixture of proteins and nucleic acids. In this review, we discuss how underlying physical and chemical processes generate internal condensate architectures. We then outline the diverse condensate architectures that are observed in biological systems. Finally, we discuss how specific condensate organization is critical for specific biological functions.
Keywords: biomolecular condensate; liquid–liquid phase separation; membraneless organelle.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources