

Identification of the Viral Determinant of Hypovirulence and Host Range in Sclerotiniaceae of a Genomovirus Reconstructed from the Plant Metagenome
- PMID: 34132570
- PMCID: PMC8354332
- DOI: 10.1128/JVI.00264-21
Identification of the Viral Determinant of Hypovirulence and Host Range in Sclerotiniaceae of a Genomovirus Reconstructed from the Plant Metagenome
Abstract
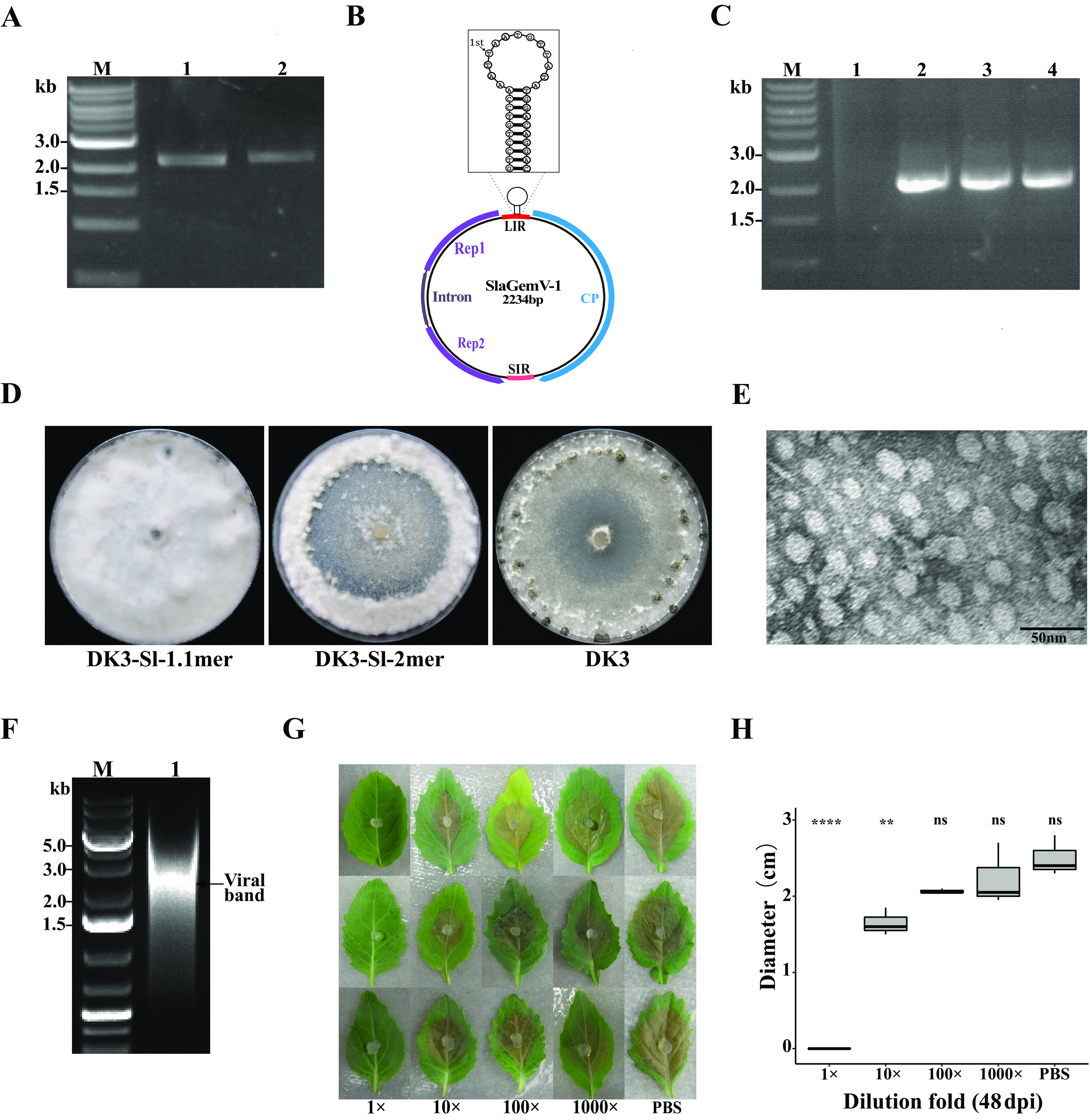
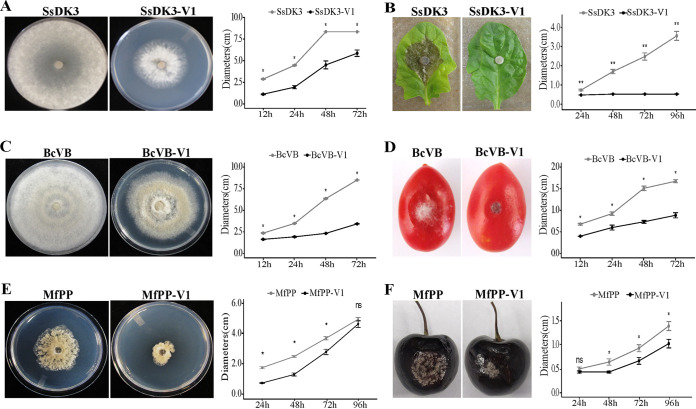
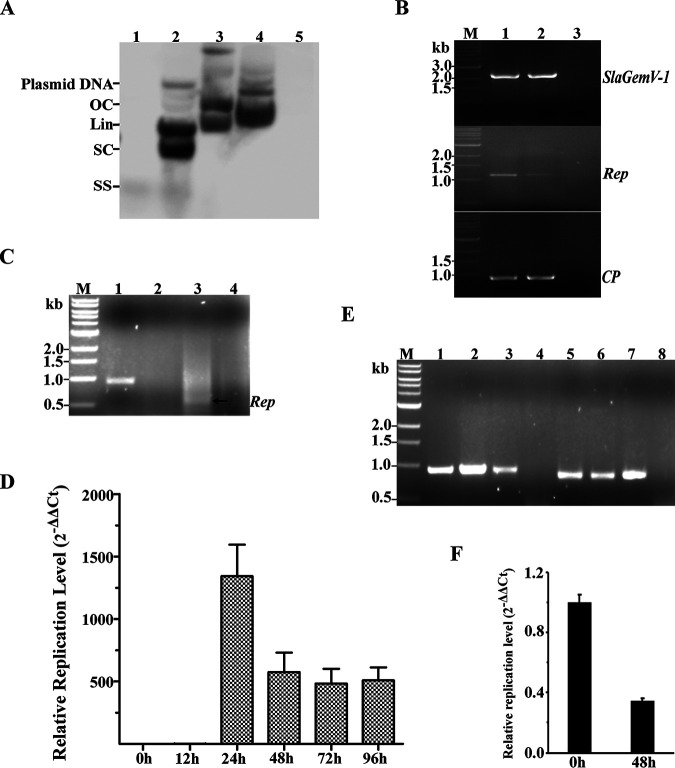
Uncharacterized viral genomes that encode circular replication-associated proteins of single-stranded DNA viruses have been discovered by metagenomics/metatranscriptomics approaches. Some of these novel viruses are classified in the newly formed family Genomoviridae. Here, we determined the host range of a novel genomovirus, SlaGemV-1, through the transfection of Sclerotinia sclerotiorum with infectious clones. Inoculating with the rescued virions, we further transfected Botrytis cinerea and Monilinia fructicola, two economically important members of the family Sclerotiniaceae, and Fusarium oxysporum. SlaGemV-1 causes hypovirulence in S. sclerotiorum, B. cinerea, and M. fructicola. SlaGemV-1 also replicates in Spodoptera frugiperda insect cells but not in Caenorhabditis elegans or plants. By expressing viral genes separately through site-specific integration, the replication protein alone was sufficient to cause debilitation. Our study is the first to demonstrate the reconstruction of a metagenomically discovered genomovirus without known hosts with the potential of inducing hypovirulence, and the infectious clone allows for studying mechanisms of genomovirus-host interactions that are conserved across genera. IMPORTANCE Little is known about the exact host range of widespread genomoviruses. The genome of soybean leaf-associated gemygorvirus-1 (SlaGemV-1) was originally assembled from a metagenomic/metatranscriptomic study without known hosts. Here, we rescued SlaGemV-1 and found that it could infect three important plant-pathogenic fungi and fall armyworm (S. frugiperda Sf9) insect cells but not a model nematode, C. elegans, or model plant species. Most importantly, SlaGemV-1 shows promise for inducing hypovirulence of the tested fungal species in the family Sclerotiniaceae, including Sclerotinia sclerotiorum, Botrytis cinerea, and Monilinia fructicola. The viral determinant of hypovirulence was further identified as replication initiation protein. As a proof of concept, we demonstrate that viromes discovered in plant metagenomes can be a valuable genetic resource when novel viruses are rescued and characterized for their host range.
Keywords: Botrytis cinerea; Sclerotinia sclerotiorum; biocontrol; gemygorvirus; genomovirus; mycovirus; virulence determinants.
Figures




Similar articles
-
Characterization of Transcriptional Responses to Genomovirus Infection of the White Mold Fungus, Sclerotinia sclerotiorum.Viruses. 2022 Aug 27;14(9):1892. doi: 10.3390/v14091892. Viruses. 2022. PMID: 36146699 Free PMC article.
-
A novel partitivirus that confers hypovirulence on plant pathogenic fungi.J Virol. 2014 Sep 1;88(17):10120-33. doi: 10.1128/JVI.01036-14. Epub 2014 Jun 25. J Virol. 2014. PMID: 24965462 Free PMC article.
-
A Mechanically Transmitted DNA Mycovirus Is Targeted by the Defence Machinery of Its Host, Botrytis cinerea.Viruses. 2021 Jul 7;13(7):1315. doi: 10.3390/v13071315. Viruses. 2021. PMID: 34372522 Free PMC article.
-
Viruses of the plant pathogenic fungus Sclerotinia sclerotiorum.Adv Virus Res. 2013;86:215-48. doi: 10.1016/B978-0-12-394315-6.00008-8. Adv Virus Res. 2013. PMID: 23498908 Review.
-
Viruses of plant-pathogenic fungi: a promising biocontrol strategy for Sclerotinia sclerotiorum.Arch Microbiol. 2023 Dec 24;206(1):38. doi: 10.1007/s00203-023-03774-8. Arch Microbiol. 2023. PMID: 38142438 Review.
Cited by
-
Characterization of Transcriptional Responses to Genomovirus Infection of the White Mold Fungus, Sclerotinia sclerotiorum.Viruses. 2022 Aug 27;14(9):1892. doi: 10.3390/v14091892. Viruses. 2022. PMID: 36146699 Free PMC article.
-
Direct Metatranscriptomic Survey of the Sunflower Microbiome and Virome.Viruses. 2021 Sep 18;13(9):1867. doi: 10.3390/v13091867. Viruses. 2021. PMID: 34578448 Free PMC article.
-
Novel Virus Identification through Metagenomics: A Systematic Review.Life (Basel). 2022 Dec 7;12(12):2048. doi: 10.3390/life12122048. Life (Basel). 2022. PMID: 36556413 Free PMC article. Review.
-
Mycovirus-induced hypovirulence in notorious fungi Sclerotinia: a comprehensive review.Braz J Microbiol. 2023 Sep;54(3):1459-1478. doi: 10.1007/s42770-023-01073-4. Epub 2023 Jul 31. Braz J Microbiol. 2023. PMID: 37523037 Free PMC article. Review.
-
Replication of single viruses across the kingdoms, Fungi, Plantae, and Animalia.Proc Natl Acad Sci U S A. 2024 Jun 18;121(25):e2318150121. doi: 10.1073/pnas.2318150121. Epub 2024 Jun 12. Proc Natl Acad Sci U S A. 2024. PMID: 38865269 Free PMC article.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
