

Antibody-Based Inhibition of Pathogenic New World Hemorrhagic Fever Mammarenaviruses by Steric Occlusion of the Human Transferrin Receptor 1 Apical Domain
- PMID: 34132574
- PMCID: PMC8354235
- DOI: 10.1128/JVI.01868-20
Antibody-Based Inhibition of Pathogenic New World Hemorrhagic Fever Mammarenaviruses by Steric Occlusion of the Human Transferrin Receptor 1 Apical Domain
Erratum in
-
Correction for Ferrero et al., "Antibody-Based Inhibition of Pathogenic New World Hemorrhagic Fever Mammarenaviruses by Steric Occlusion of the Human Transferrin Receptor 1 Apical Domain".J Virol. 2022 Feb 23;96(4):e0216321. doi: 10.1128/jvi.02163-21. Epub 2022 Feb 23. J Virol. 2022. PMID: 35195450 Free PMC article. No abstract available.
Abstract
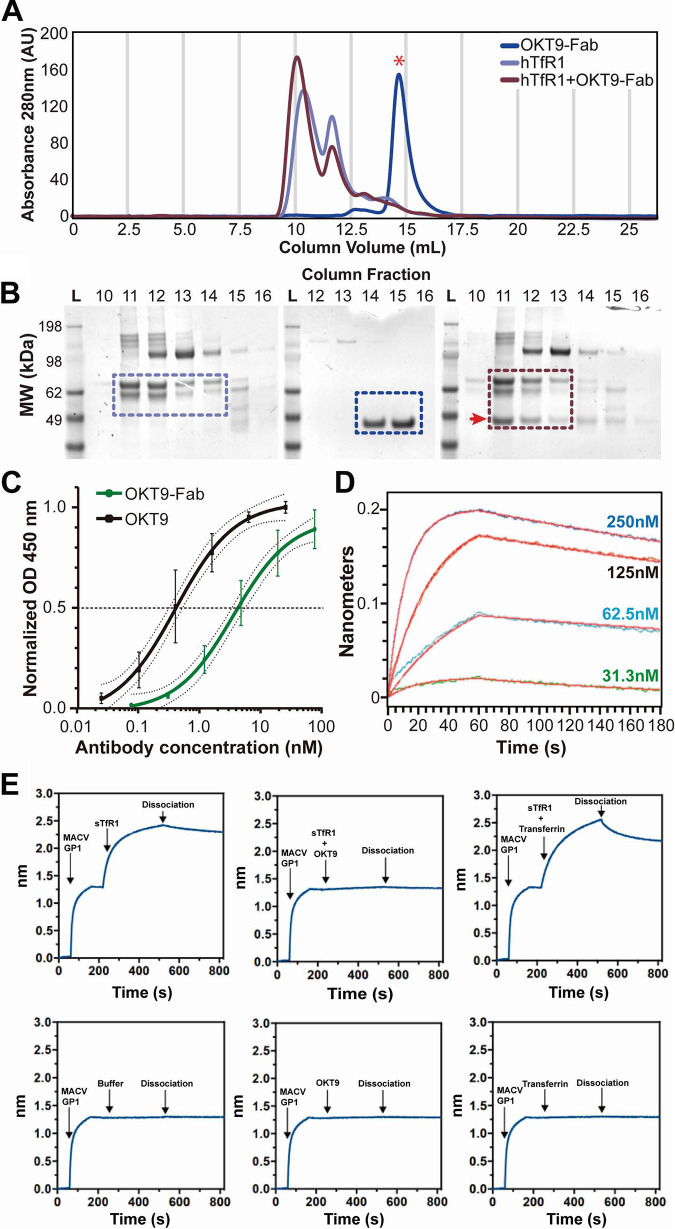
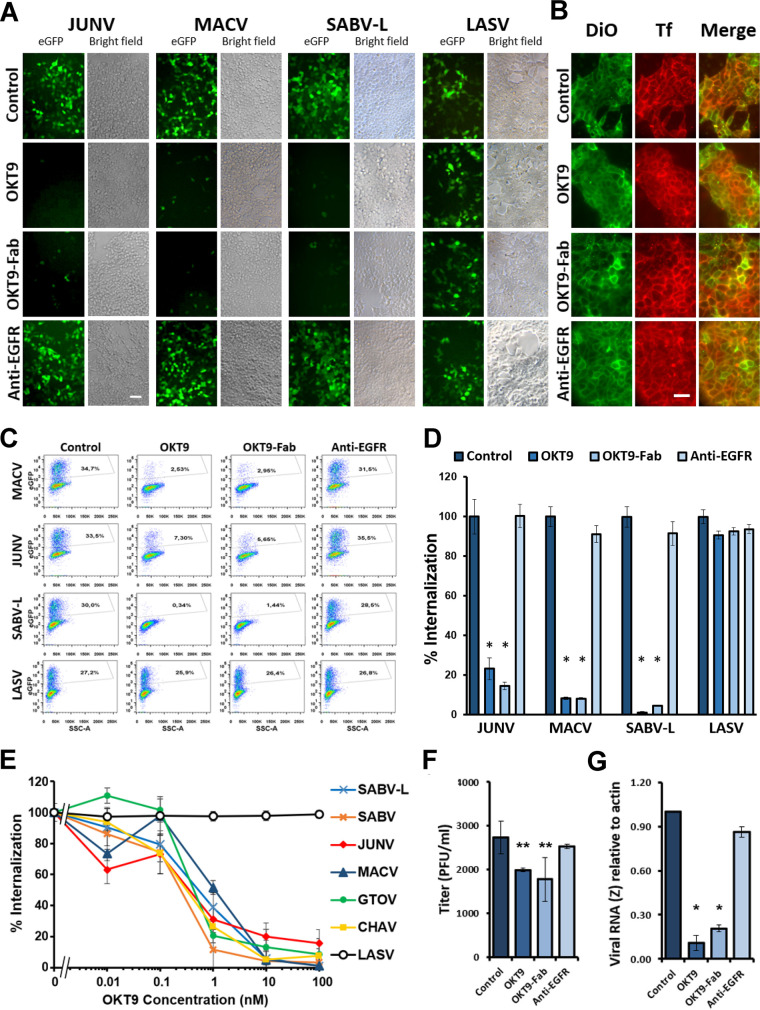
Pathogenic clade B New World mammarenaviruses (NWM) can cause Argentine, Venezuelan, Brazilian, and Bolivian hemorrhagic fevers. Sequence variability among NWM glycoproteins (GP) poses a challenge to the development of broadly neutralizing therapeutics against the entire clade of viruses. However, blockade of their shared binding site on the apical domain of human transferrin receptor 1 (hTfR1/CD71) presents an opportunity for the development of effective and broadly neutralizing therapeutics. Here, we demonstrate that the murine monoclonal antibody OKT9, which targets the apical domain of hTfR1, can sterically block cellular entry by viral particles presenting clade B NWM glycoproteins (GP1-GP2). OKT9 blockade is also effective against viral particles pseudotyped with glycoproteins of a recently identified pathogenic Sabia-like virus. With nanomolar affinity for hTfR1, the OKT9 antigen binding fragment (OKT9-Fab) sterically blocks clade B NWM-GP1s and reduces infectivity of an attenuated strain of Junin virus. Binding of OKT9 to the hTfR1 ectodomain in its soluble, dimeric state produces stable assemblies that are observable by negative-stain electron microscopy. A model of the OKT9-sTfR1 complex, informed by the known crystallographic structure of sTfR1 and a newly determined structure of the OKT9 antigen binding fragment (Fab), suggests that OKT9 and the Machupo virus GP1 share a binding site on the hTfR1 apical domain. The structural basis for this interaction presents a framework for the design and development of high-affinity, broadly acting agents targeting clade B NWMs. IMPORTANCE Pathogenic clade B NWMs cause grave infectious diseases, the South American hemorrhagic fevers. Their etiological agents are Junin (JUNV), Guanarito (GTOV), Sabiá (SABV), Machupo (MACV), Chapare (CHAV), and a new Sabiá-like (SABV-L) virus recently identified in Brazil. These are priority A pathogens due to their high infectivity and mortality, their potential for person-to-person transmission, and the limited availability of effective therapeutics and vaccines to curb their effects. While low homology between surface glycoproteins of NWMs foils efforts to develop broadly neutralizing therapies targeting NWMs, this work provides structural evidence that OKT9, a monoclonal antibody targeting a single NWM glycoprotein binding site on hTfR1, can efficiently prevent their entry into cells.
Keywords: Junin; Machupo; Sabiá-like; X-ray crystallography; antiviral agents; electron microscopy; mammarenavirus; monoclonal antibodies; transferrin receptor.
Figures
Similar articles
-
Host receptor-targeted therapeutic approach to counter pathogenic New World mammarenavirus infections.Nat Commun. 2022 Jan 28;13(1):558. doi: 10.1038/s41467-021-27949-3. Nat Commun. 2022. PMID: 35091550 Free PMC article.
-
Glycoprotein-Specific Antibodies Produced by DNA Vaccination Protect Guinea Pigs from Lethal Argentine and Venezuelan Hemorrhagic Fever.J Virol. 2016 Jan 20;90(7):3515-29. doi: 10.1128/JVI.02969-15. J Virol. 2016. PMID: 26792737 Free PMC article.
-
A hTfR1 Receptor-Specific VHH Antibody Neutralizes Pseudoviruses Expressing Glycoproteins from Junín and Machupo Viruses.Viruses. 2024 Dec 20;16(12):1951. doi: 10.3390/v16121951. Viruses. 2024. PMID: 39772257 Free PMC article.
-
Drug discovery technologies and strategies for Machupo virus and other New World arenaviruses.Expert Opin Drug Discov. 2012 Jul;7(7):613-32. doi: 10.1517/17460441.2012.687719. Epub 2012 May 19. Expert Opin Drug Discov. 2012. PMID: 22607481 Free PMC article. Review.
-
Structural and molecular biology of Sabiá virus.Exp Biol Med (Maywood). 2023 Oct;248(19):1624-1634. doi: 10.1177/15353702231199071. Epub 2023 Nov 8. Exp Biol Med (Maywood). 2023. PMID: 37937408 Free PMC article. Review.
Cited by
-
Host receptor-targeted therapeutic approach to counter pathogenic New World mammarenavirus infections.Nat Commun. 2022 Jan 28;13(1):558. doi: 10.1038/s41467-021-27949-3. Nat Commun. 2022. PMID: 35091550 Free PMC article.
-
An AAV capsid reprogrammed to bind human Transferrin Receptor mediates brain-wide gene delivery.bioRxiv [Preprint]. 2023 Dec 22:2023.12.20.572615. doi: 10.1101/2023.12.20.572615. bioRxiv. 2023. Update in: Science. 2024 Jun 14;384(6701):1220-1227. doi: 10.1126/science.adm8386. PMID: 38187643 Free PMC article. Updated. Preprint.
-
Repurposing an endogenous degradation domain for antibody-mediated disposal of cell-surface proteins.EMBO Rep. 2024 Mar;25(3):951-970. doi: 10.1038/s44319-024-00063-3. Epub 2024 Jan 29. EMBO Rep. 2024. PMID: 38287192 Free PMC article.
-
Temporal changes in pathology and viral RNA distribution in guinea pigs following separate infection with two New World Arenaviruses.PLoS Negl Trop Dis. 2023 Sep 8;17(9):e0011620. doi: 10.1371/journal.pntd.0011620. eCollection 2023 Sep. PLoS Negl Trop Dis. 2023. PMID: 37682988 Free PMC article.
-
Synthesis and Evaluation of [64Cu]Cu-NOTA-HFn for PET Imaging of Transferrin Receptor 1 Expression in Nasopharyngeal Carcinoma.ACS Omega. 2024 Apr 5;9(15):17423-17431. doi: 10.1021/acsomega.4c00187. eCollection 2024 Apr 16. ACS Omega. 2024. PMID: 38645324 Free PMC article.
References
-
- Borio L, Inglesby T, Peters CJ, Schmaljohn AL, Hughes JM, Jahrling PB, Ksiazek T, Johnson KM, Meyerhoff A, O'Toole T, Ascher MS, Bartlett J, Breman JG, Eitzen EM, Jr, Hamburg M, Hauer J, Henderson DA, Johnson RT, Kwik G, Layton M, Lillibridge S, Nabel GJ, Osterholm MT, Perl TM, Russell P, Tonat K, Working Group on Civilian Biodefense. 2002. Hemorrhagic fever viruses as biological weapons: medical and public health management. JAMA 287:2391–2405. 10.1001/jama.287.18.2391. - DOI - PubMed
-
- Centers for Disease Control and Prevention. 2012. Possession, use, and transfer of select agents and toxins; biennial review. Final rule. Fed Regist 77:61083–61115. - PubMed
-
- Zeitlin L, Geisbert JB, Deer DJ, Fenton KA, Bohorov O, Bohorova N, Goodman C, Kim D, Hiatt A, Pauly MH, Velasco J, Whaley KJ, Altmann F, Gruber C, Steinkellner H, Honko AN, Kuehne AI, Aman MJ, Sahandi S, Enterlein S, Zhan X, Enria D, Geisbert TW. 2016. Monoclonal antibody therapy for Junin virus infection. Proc Natl Acad Sci U S A 113:4458–4463. 10.1073/pnas.1600996113. - DOI - PMC - PubMed
-
- Westover JB, Sefing EJ, Bailey KW, Van Wettere AJ, Jung KH, Dagley A, Wandersee L, Downs B, Smee DF, Furuta Y, Bray M, Gowen BB. 2016. Low-dose ribavirin potentiates the antiviral activity of favipiravir against hemorrhagic fever viruses. Antiviral Res 126:62–68. 10.1016/j.antiviral.2015.12.006. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- DE-FC02-02ER63421/U.S. Department of Energy (DOE)
- P41 GM136508/GM/NIGMS NIH HHS/United States
- T32 GM007185/GM/NIGMS NIH HHS/United States
- IP-COVID19-929/MINCyT | ANPCyT | Fondo para la Investigación Científica y Tecnológica (FonCyT)
- R35 GM128867/GM/NIGMS NIH HHS/United States
- DMR-1548924/National Science Foundation (NSF)
- R21 AI135851/AI/NIAID NIH HHS/United States
- Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
- 5R21AI135851-02/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- PICT 2018-3954/MINCyT | ANPCyT | Fondo para la Investigación Científica y Tecnológica (FonCyT)
- P30 GM124165/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
