

Sneezing reflex is mediated by a peptidergic pathway from nose to brainstem
- PMID: 34133943
- PMCID: PMC8396370
- DOI: 10.1016/j.cell.2021.05.017
Sneezing reflex is mediated by a peptidergic pathway from nose to brainstem
Abstract
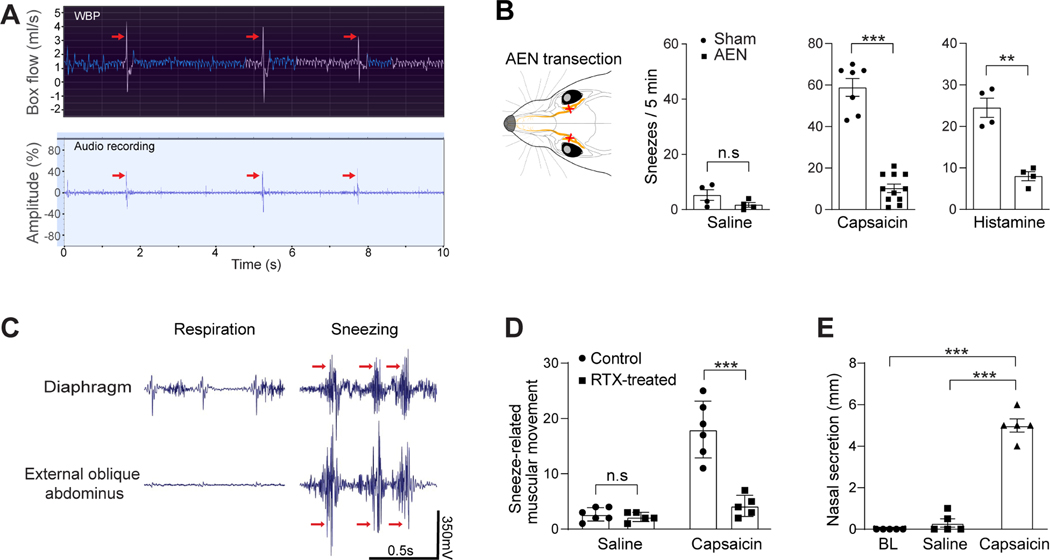
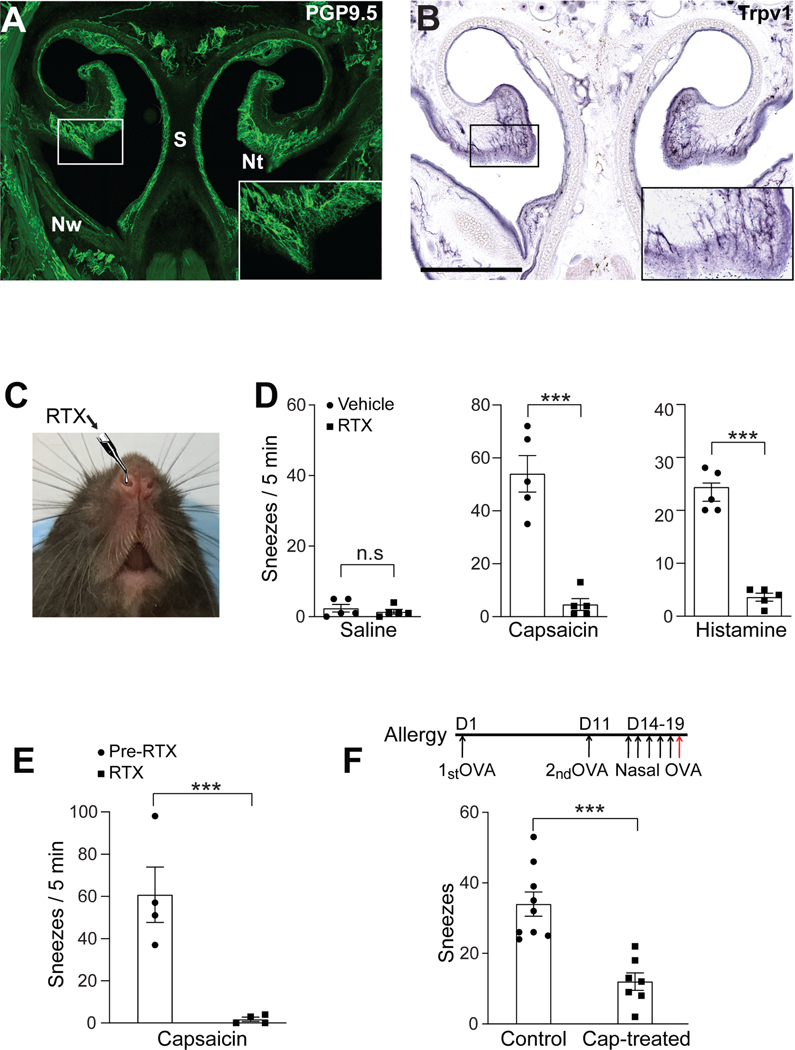
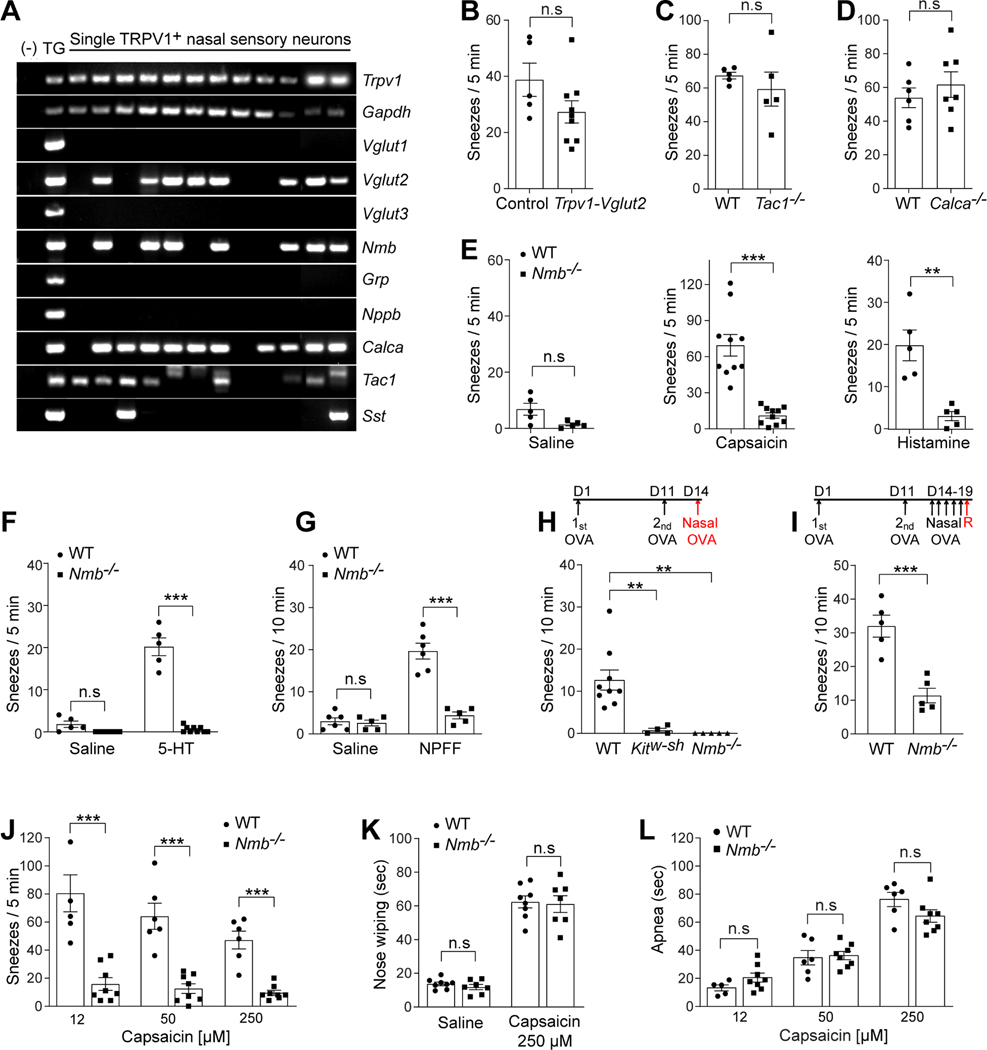
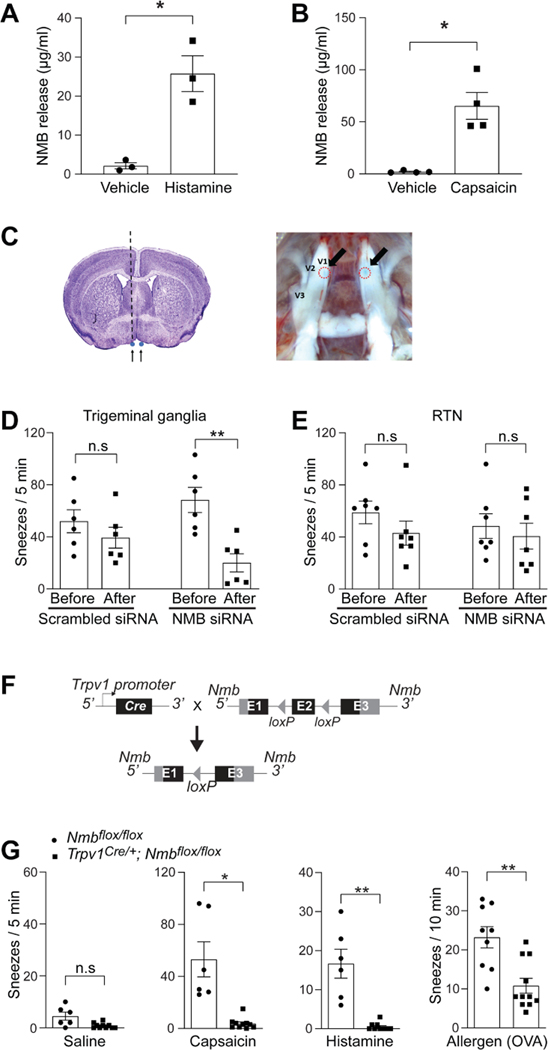
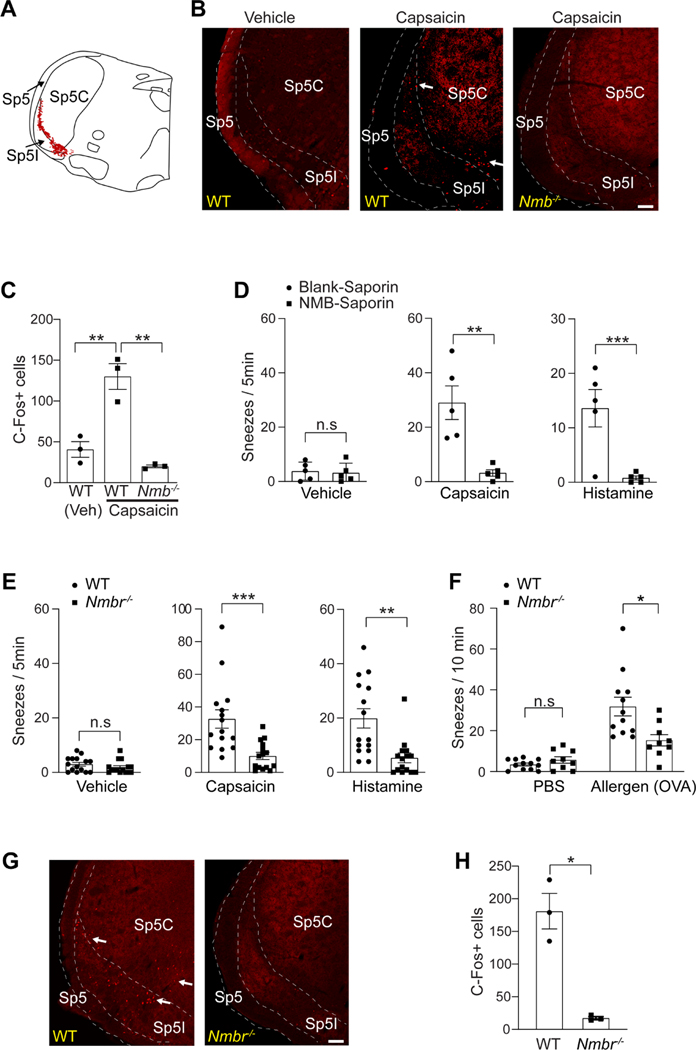
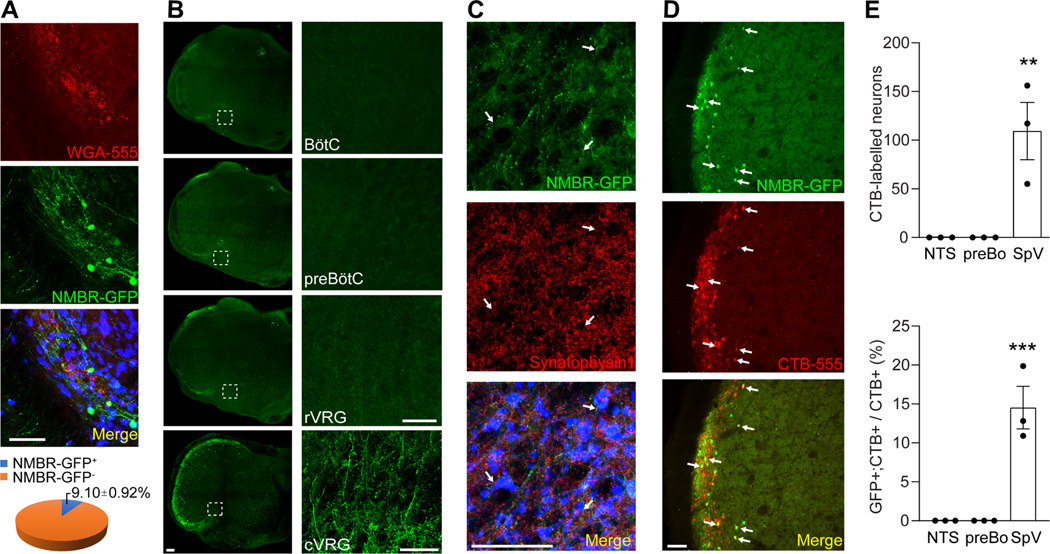
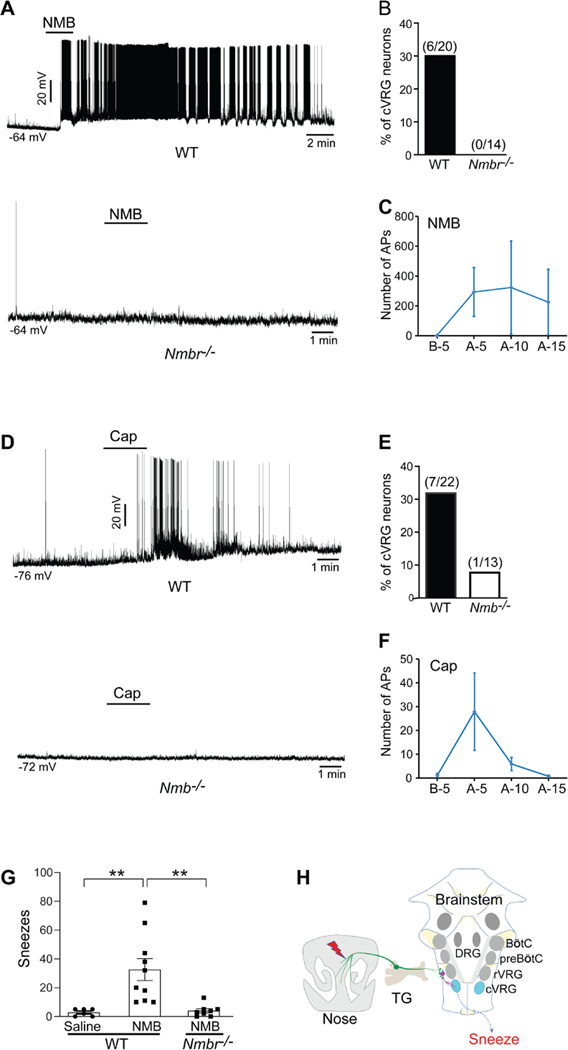
Sneezing is a vital respiratory reflex frequently associated with allergic rhinitis and viral respiratory infections. However, its neural circuit remains largely unknown. A sneeze-evoking region was discovered in both cat and human brainstems, corresponding anatomically to the central recipient zone of nasal sensory neurons. Therefore, we hypothesized that a neuronal population postsynaptic to nasal sensory neurons mediates sneezing in this region. By screening major presynaptic neurotransmitters/neuropeptides released by nasal sensory neurons, we found that neuromedin B (NMB) peptide is essential for signaling sneezing. Ablation of NMB-sensitive postsynaptic neurons in the sneeze-evoking region or deficiency in NMB receptor abolished the sneezing reflex. Remarkably, NMB-sensitive neurons further project to the caudal ventral respiratory group (cVRG). Chemical activation of NMB-sensitive neurons elicits action potentials in cVRG neurons and leads to sneezing behavior. Our study delineates a peptidergic pathway mediating sneezing, providing molecular insights into the sneezing reflex arc.
Keywords: caudal ventral respiratory group; nasal sensory neurons; neuropeptide; sneeze; sneeze-evoking region.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests. B.S.K. has noncompeting financial interests unrelated to this work and has served as a consultant for AbbVie, ABRAX Japan, Almirall, Cara Therapeutics, Maruho, Menlo Therapeutics, Pfizer, and Third Rock Ventures. He has participated on the advisory board for Almirall, Boehringer Ingelheim, Cara Therapeutics, Kiniksa Pharmaceuticals, Menlo Therapeutics, Regeneron Pharmaceuticals, Sanofi Genzyme, and Trevi Therapeutics. He is also founder, chief scientific officer, and stockholder of Nuogen Pharma and stockholder of Locus Biosciences.
Figures
Comment in
-
Sneezing pathway revealed.Nat Rev Neurosci. 2021 Aug;22(8):455. doi: 10.1038/s41583-021-00491-3. Nat Rev Neurosci. 2021. PMID: 34226717 No abstract available.
References
-
- Batsel HL, and Lines AJ (1978). Discharge of respiratory neurons in sneezes resulting from ethmoidal nerve stimulation. Exp Neurol 58, 410–424. - PubMed
-
- Bourouiba L. (2016). IMAGES IN CLINICAL MEDICINE. A Sneeze. N Engl J Med 375, e15. - PubMed
-
- Bourouiba L. (2020). Turbulent Gas Clouds and Respiratory Pathogen Emissions: Potential Implications for Reducing Transmission of COVID-19. JAMA 323, 1837–1838. - PubMed
-
- Bourouiba L, Dehandschoewercker E, and M.B.J.W. (2014). Violent expiratory events: on coughing and sneezing. J Fluid Mech 745, 27.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
