

The role of pectin phase separation in plant cell wall assembly and growth
- PMID: 34141960
- PMCID: PMC8185244
- DOI: 10.1016/j.tcsw.2021.100054
The role of pectin phase separation in plant cell wall assembly and growth
Abstract
A rapidly increasing body of literature suggests that many biological processes are driven by phase separation within polymer mixtures. Liquid-liquid phase separation can lead to the formation of membrane-less organelles, which are thought to play a wide variety of roles in cell metabolism, gene regulation or signaling. One of the characteristics of these systems is that they are poised at phase transition boundaries, which makes them perfectly suited to elicit robust cellular responses to often very small changes in the cell's "environment". Recent observations suggest that, also in the semi-solid environment of plant cell walls, phase separation not only plays a role in wall patterning, hydration and stress relaxation during growth, but also may provide a driving force for cell wall expansion. In this context, pectins, the major polyanionic polysaccharides in the walls of growing cells, appear to play a critical role. Here, we will discuss (i) our current understanding of the structure-function relationship of pectins, (ii) in vivo evidence that pectin modification can drive critical phase transitions in the cell wall, (iii) how such phase transitions may drive cell wall expansion in addition to turgor pressure and (iv) the periodic cellular processes that may control phase transitions underlying cell wall assembly and expansion.
Keywords: Cell expansion; Pectin; Phase separation; Plant cell wall; Volume transition.
© 2021 The Author(s).
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures




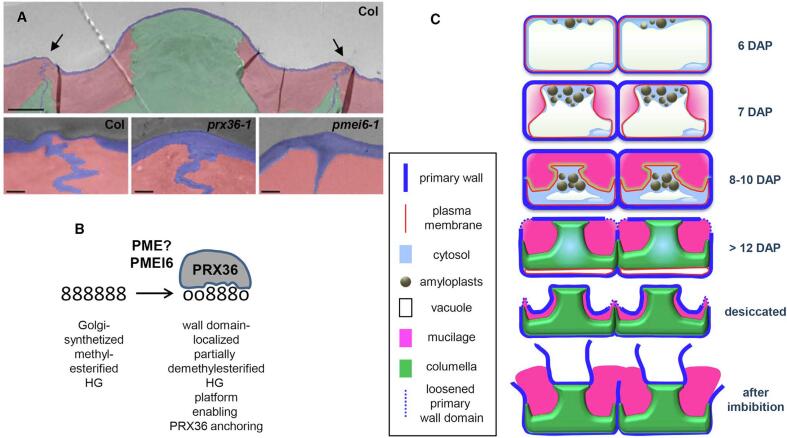

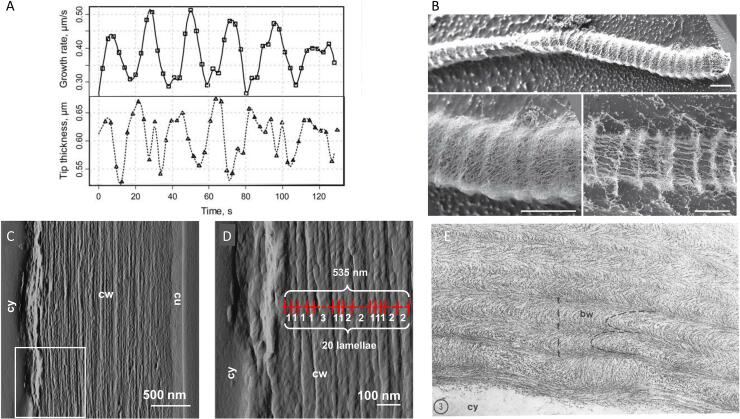
References
-
- Arend, M., Muninger, M., Fromm, J., 2008. Unique occurrence of pectin-like fibrillar cell wall deposits in xylem fibres of poplar. Plant Biol. https://doi.org/10.1111/j.1438-8677.2008.00082.x. - PubMed
-
- Baskin, T.I., 2005. Anisotropic expansion of the plant cell wall. Annu. Rev. Cell Dev. Biol. 21, 203–222. https://doi.org/https://doi.org/10.1146/annurev.cellbio.20.082503.103053. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
