

Auxin-degron system identifies immediate mechanisms of OCT4
- PMID: 34143975
- PMCID: PMC8282470
- DOI: 10.1016/j.stemcr.2021.05.016
Auxin-degron system identifies immediate mechanisms of OCT4
Abstract
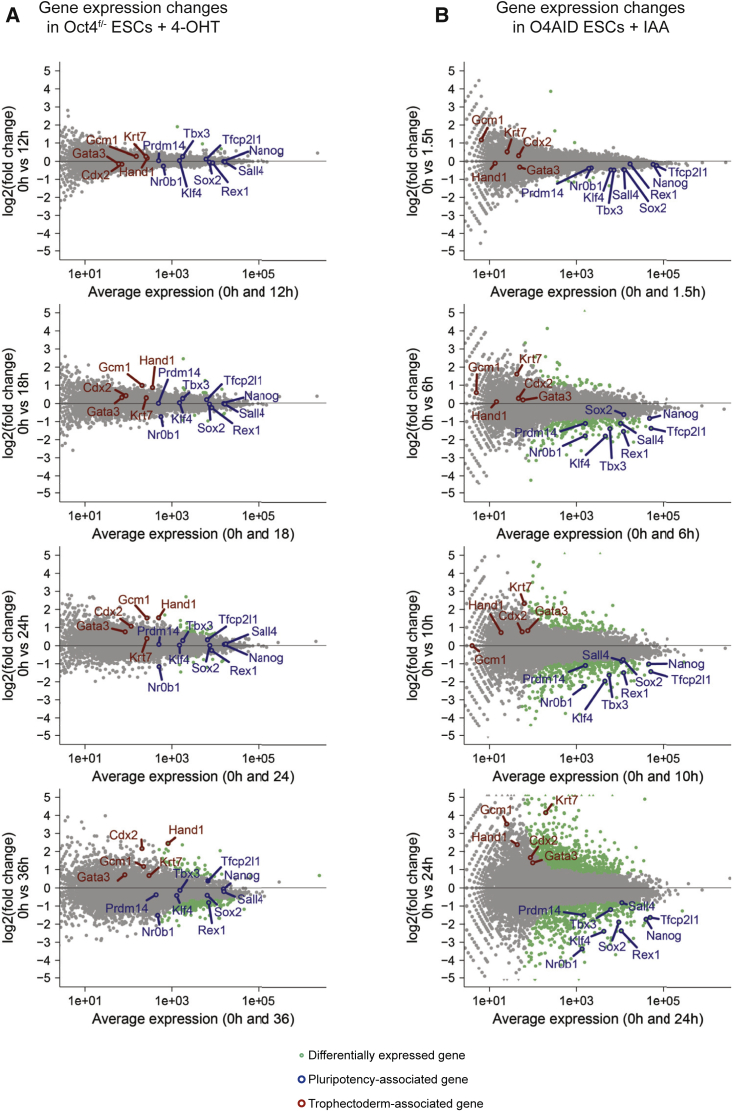
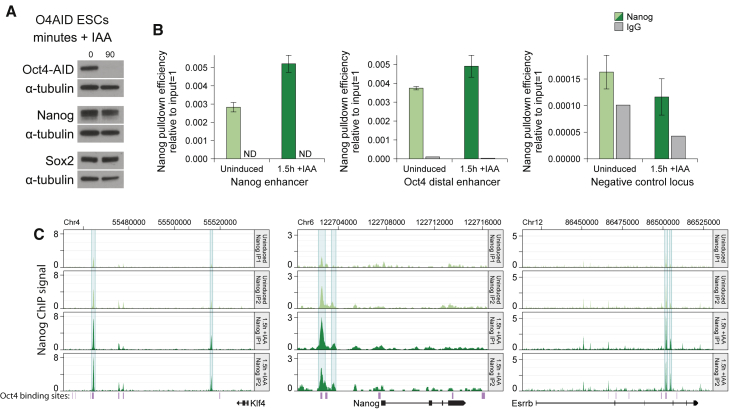
The pluripotency factor OCT4 is essential for the maintenance of naive pluripotent stem cells in vitro and in vivo. However, the specific role of OCT4 in this process remains unknown. Here, we developed a rapid protein-level OCT4 depletion system that demonstrates that the immediate downstream response to loss of OCT4 is reduced expression of key pluripotency factors. Our data show a requirement for OCT4 for the efficient transcription of several key pluripotency factors and suggest that expression of trophectoderm markers is a subsequent event. In addition, we find that NANOG is able to bind to the genome in the absence of OCT4, and this binding is in fact enhanced. Globally, however, the active enhancer-associated histone mark H3K27ac is depleted. Our work establishes that, while OCT4 is required for the maintenance of the naive transcription factor network, at a normal embryonic stem cell levels it antagonizes this network through inhibition of NANOG binding.
Keywords: Nanog; Oct4; Pou5f1; auxin-inducible degron; embryonic stem cells; mouse nPSCs.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




References
-
- Chambers I., Silva J., Colby D., Nichols J., Nijmeijer B., Robertson M., Vrana J., Jones K., Grotewold L., Smith A. Nanog safeguards pluripotency and mediates germline development. Nature. 2007;450:1230–1234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
