

Systematic profiling of protein complex dynamics reveals DNA-PK phosphorylation of IFI16 en route to herpesvirus immunity
- PMID: 34144993
- PMCID: PMC8213230
- DOI: 10.1126/sciadv.abg6680
Systematic profiling of protein complex dynamics reveals DNA-PK phosphorylation of IFI16 en route to herpesvirus immunity
Abstract
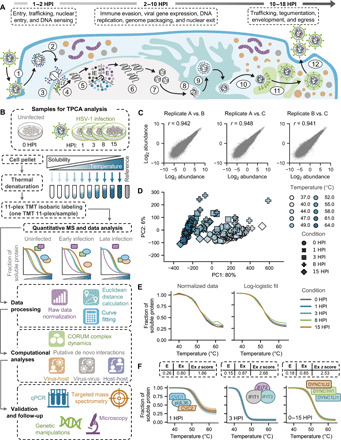
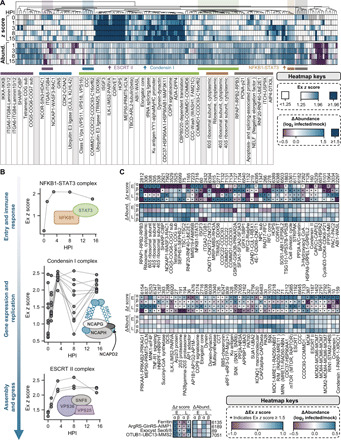
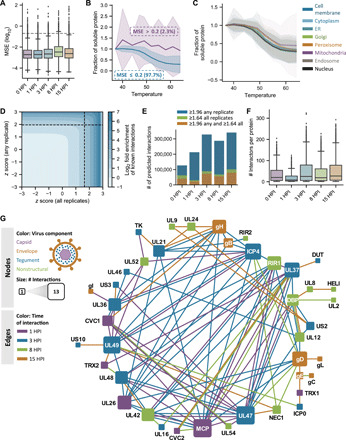
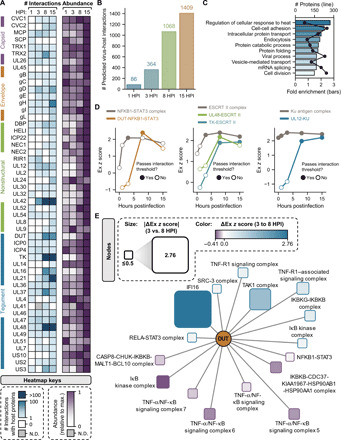
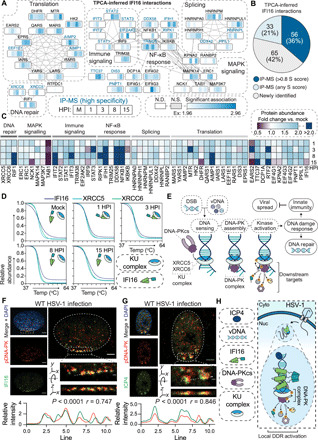
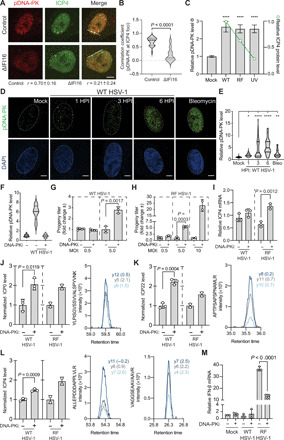
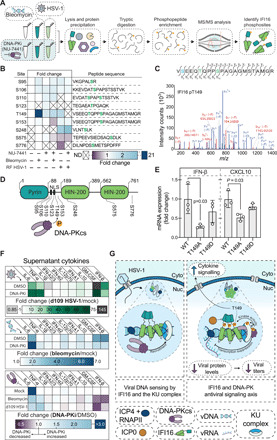
Dynamically shifting protein-protein interactions (PPIs) regulate cellular responses to viruses and the resulting immune signaling. Here, we use thermal proximity coaggregation (TPCA) mass spectrometry to characterize the on-off behavior of PPIs during infection with herpes simplex virus 1 (HSV-1), a virus with an ancient history of coevolution with hosts. Advancing the TPCA analysis to infer associations de novo, we build a time-resolved portrait of thousands of host-host, virus-host, and virus-virus PPIs. We demonstrate that, early in infection, the DNA sensor IFI16 recruits the active DNA damage response kinase, DNA-dependent protein kinase (DNA-PK), to incoming viral DNA at the nuclear periphery. We establish IFI16 T149 as a substrate of DNA-PK upon viral infection or DNA damage. This phosphorylation promotes IFI16-driven cytokine responses. Together, we characterize the global dynamics of PPIs during HSV-1 infection, uncovering the co-regulation of IFI16 and DNA-PK functions as a missing link in immunity to herpesvirus infection.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Whitley R. J., Roizman B., Herpes simplex virus infections. Lancet 357, 1513–1518 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
