

Niche adaptation promoted the evolutionary diversification of tiny ocean predators
- PMID: 34155140
- PMCID: PMC8237690
- DOI: 10.1073/pnas.2020955118
Niche adaptation promoted the evolutionary diversification of tiny ocean predators
Abstract
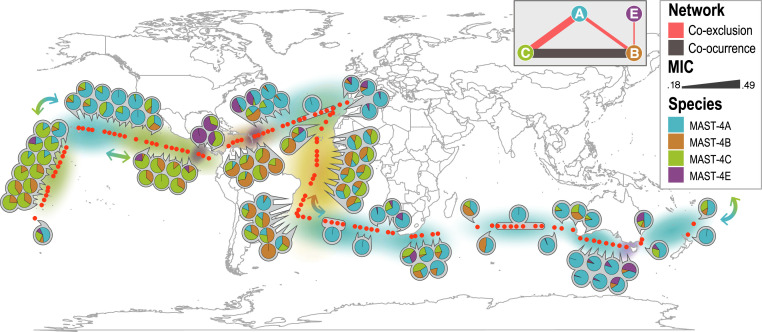
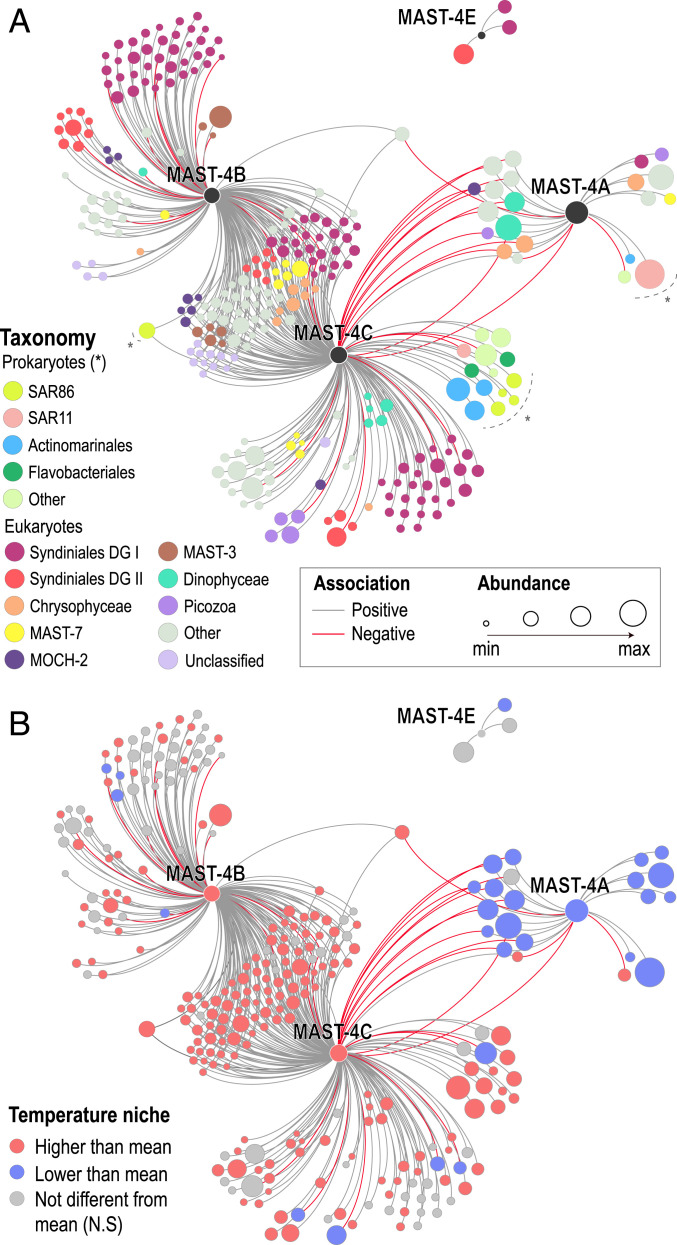
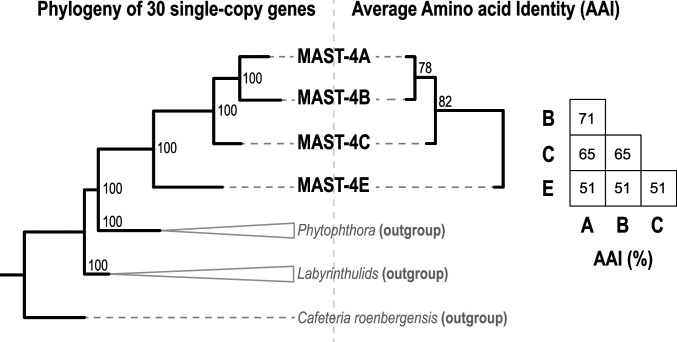
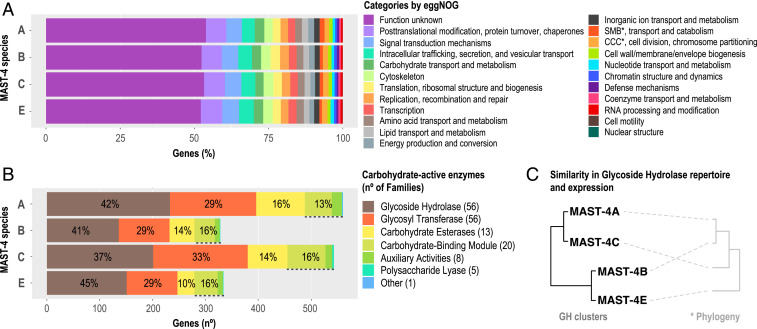
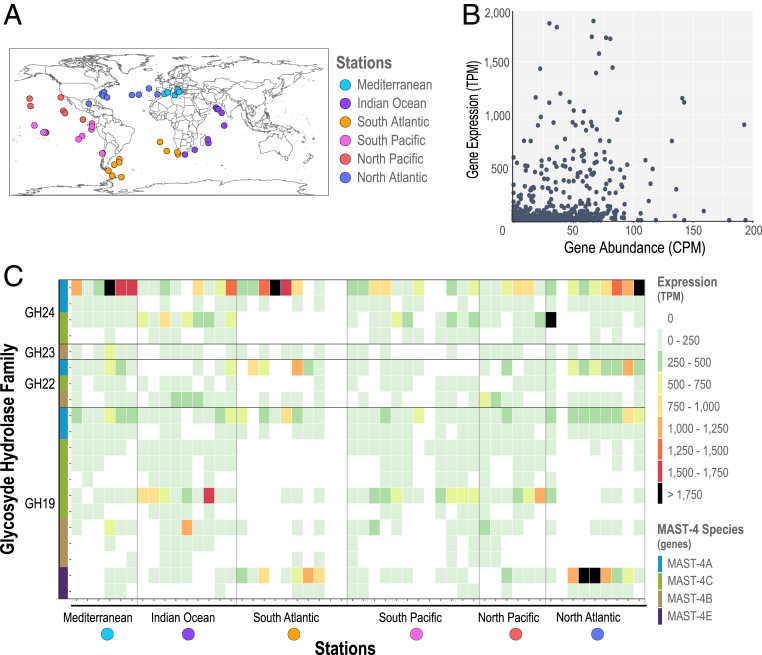
Unicellular eukaryotic predators play a crucial role in the functioning of the ocean ecosystem by recycling nutrients and energy that are channeled to upper trophic levels. Traditionally, these evolutionarily diverse organisms have been combined into a single functional group (heterotrophic flagellates), overlooking their organismal differences. Here, we investigated four evolutionarily related species belonging to one cosmopolitan group of uncultured marine picoeukaryotic predators: marine stramenopiles (MAST)-4 (species A, B, C, and E). Co-occurrence and distribution analyses in the global surface ocean indicated contrasting patterns in MAST-4A and C, suggesting adaptation to different temperatures. We then investigated whether these spatial distribution patterns were mirrored by MAST-4 genomic content using single-cell genomics. Analyses of 69 single cells recovered 66 to 83% of the MAST-4A/B/C/E genomes, which displayed substantial interspecies divergence. MAST-4 genomes were similar in terms of broad gene functional categories, but they differed in enzymes of ecological relevance, such as glycoside hydrolases (GHs), which are part of the food degradation machinery in MAST-4. Interestingly, MAST-4 species featuring a similar GH composition (A and C) coexcluded each other in the surface global ocean, while species with a different set of GHs (B and C) appeared to be able to coexist, suggesting further niche diversification associated with prey digestion. We propose that differential niche adaptation to temperature and prey type has promoted adaptive evolutionary diversification in MAST-4. We show that minute ocean predators from the same phylogenetic group may have different biogeography and genomic content, which needs to be accounted for to better comprehend marine food webs.
Keywords: MAST-4; biogeography; ecoevolution; phagocytosis; protists.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Field C. B., Behrenfeld M. J., Randerson J. T., Falkowski P., Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 281, 237–240 (1998). - PubMed
-
- Li W. K. W., Primary production of prochlorophytes, cyanobacteria, and eucaryotic ultraphytoplankton: Measurements from flow cytometric sorting. Limnol. Oceanogr. 39, 169–175 (1994).
-
- Worden A. Z., et al., Rethinking the marine carbon cycle: Factoring in the multifarious lifestyles of microbes. Science 347, 1257594 (2015). - PubMed
-
- Pernthaler J., Predation on prokaryotes in the water column and its ecological implications. Nat. Rev. Microbiol. 3, 537–546 (2005). - PubMed
-
- Massana R., et al., Grazing rates and functional diversity of uncultured heterotrophic flagellates. ISME J. 3, 588–596 (2009). - PubMed
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
