

Integrated genomic, epidemiologic investigation of Candida auris skin colonization in a skilled nursing facility
- PMID: 34155414
- PMCID: PMC9396956
- DOI: 10.1038/s41591-021-01383-w
Integrated genomic, epidemiologic investigation of Candida auris skin colonization in a skilled nursing facility
Abstract
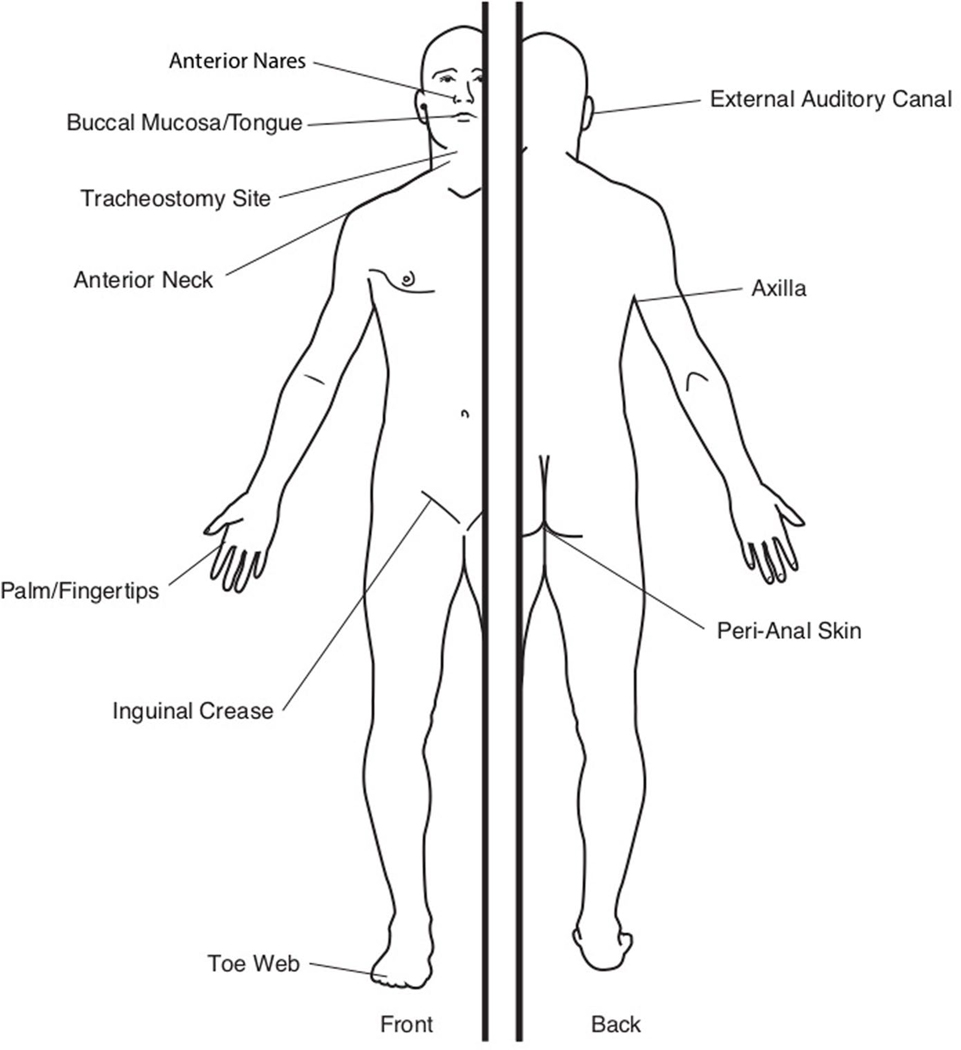
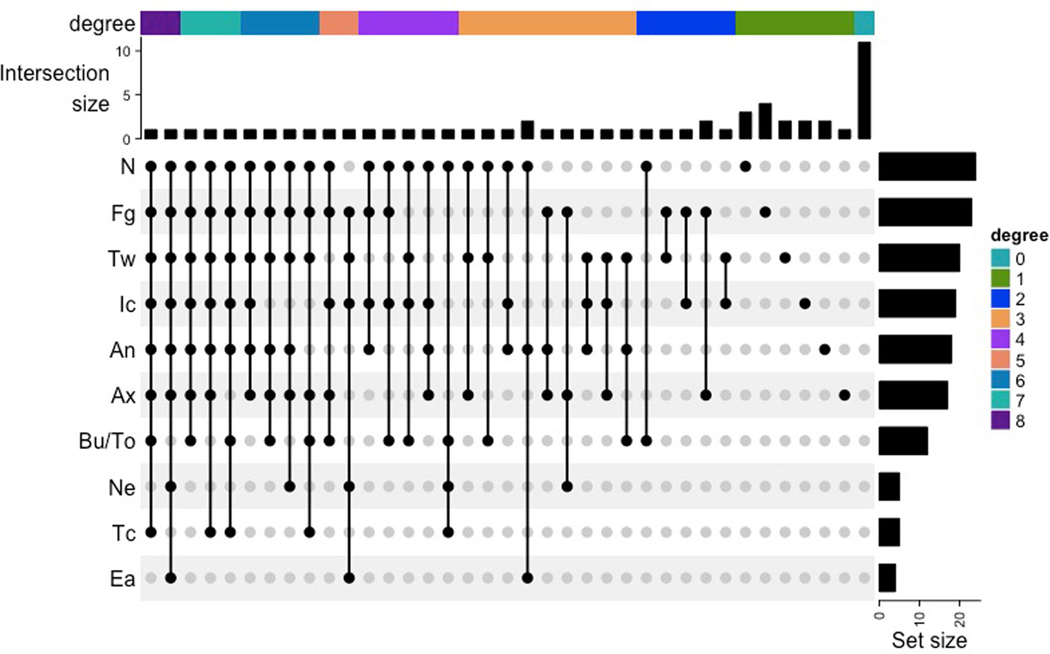
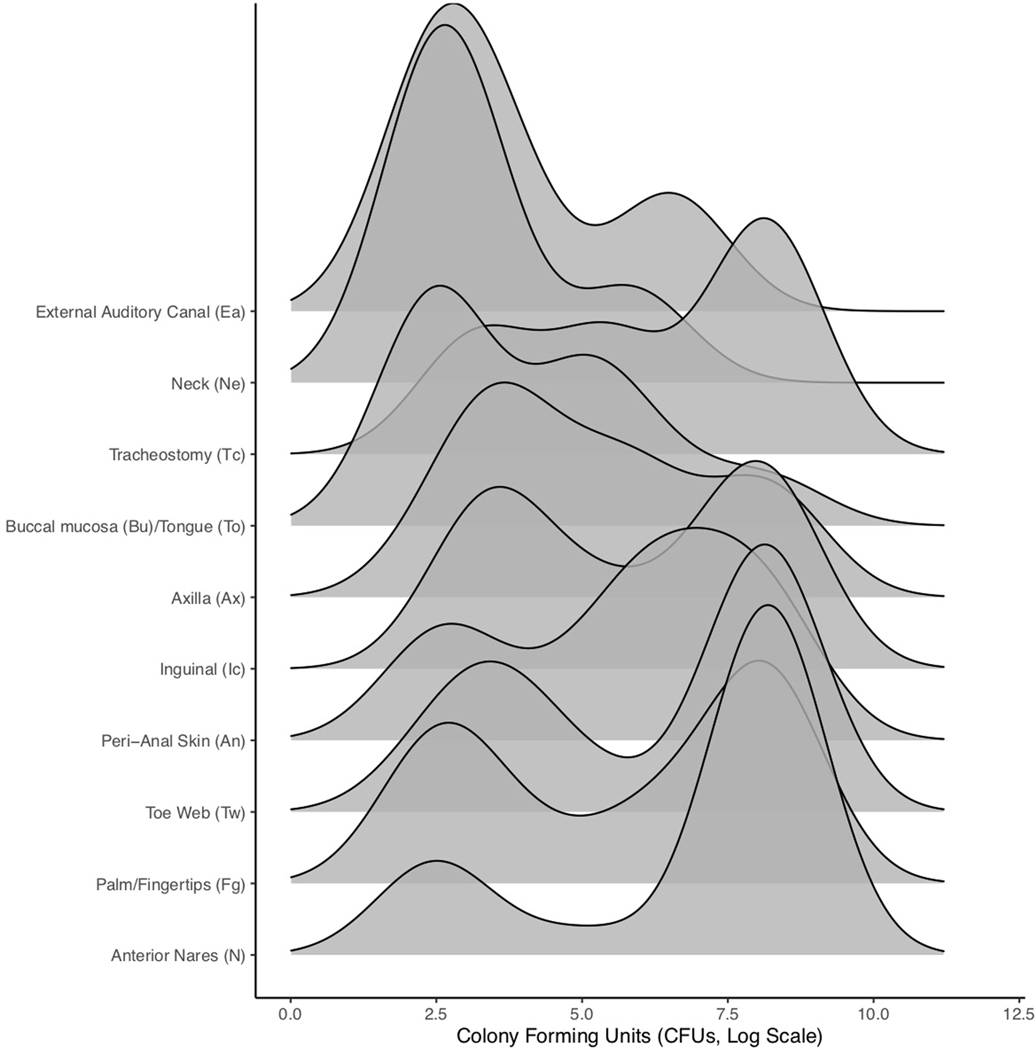
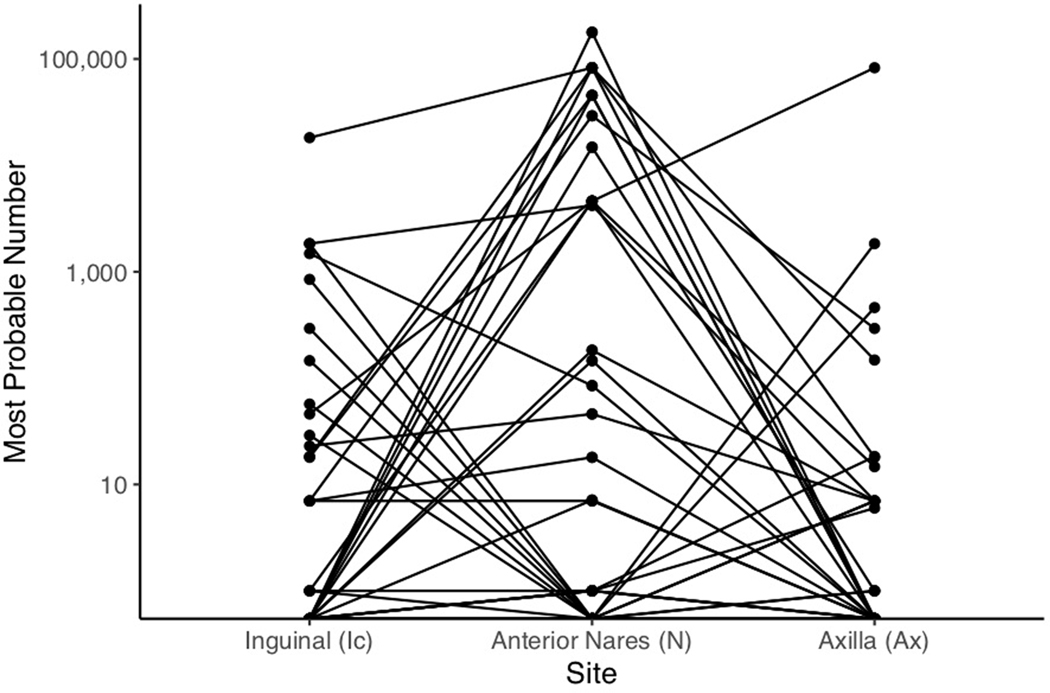
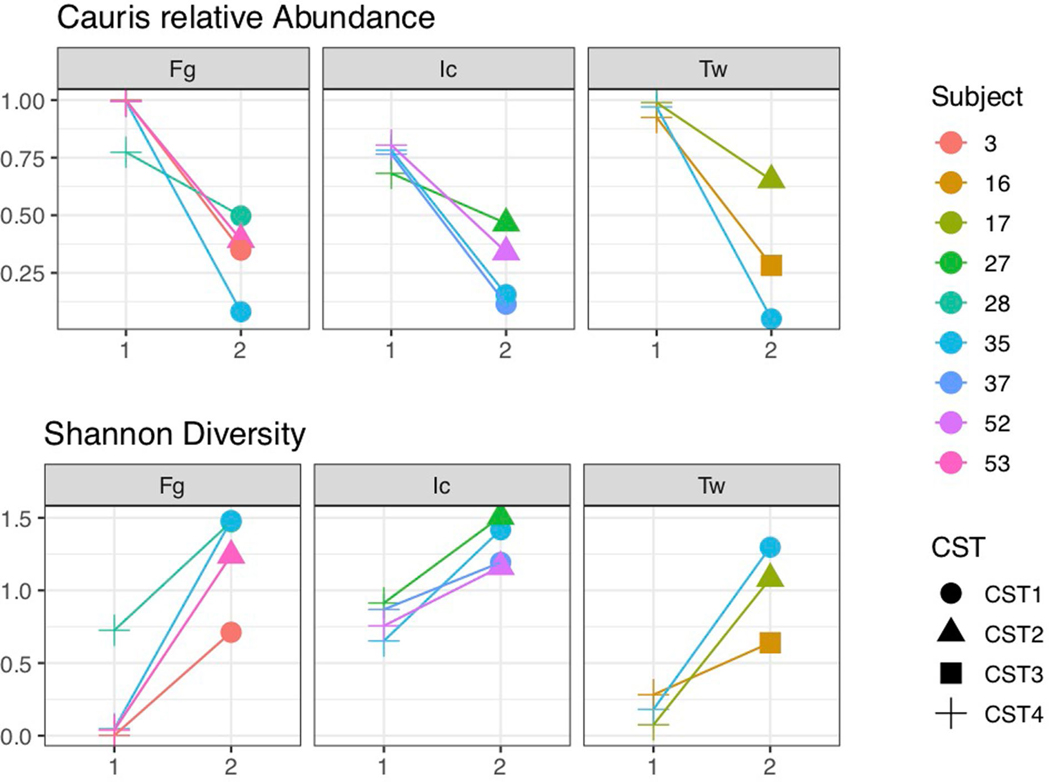
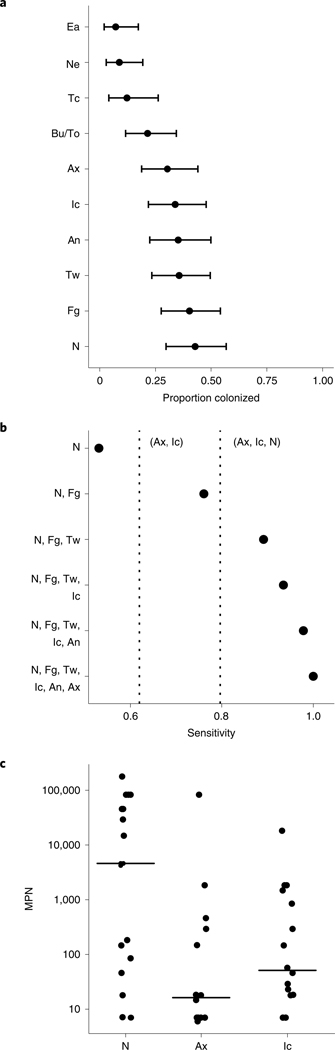
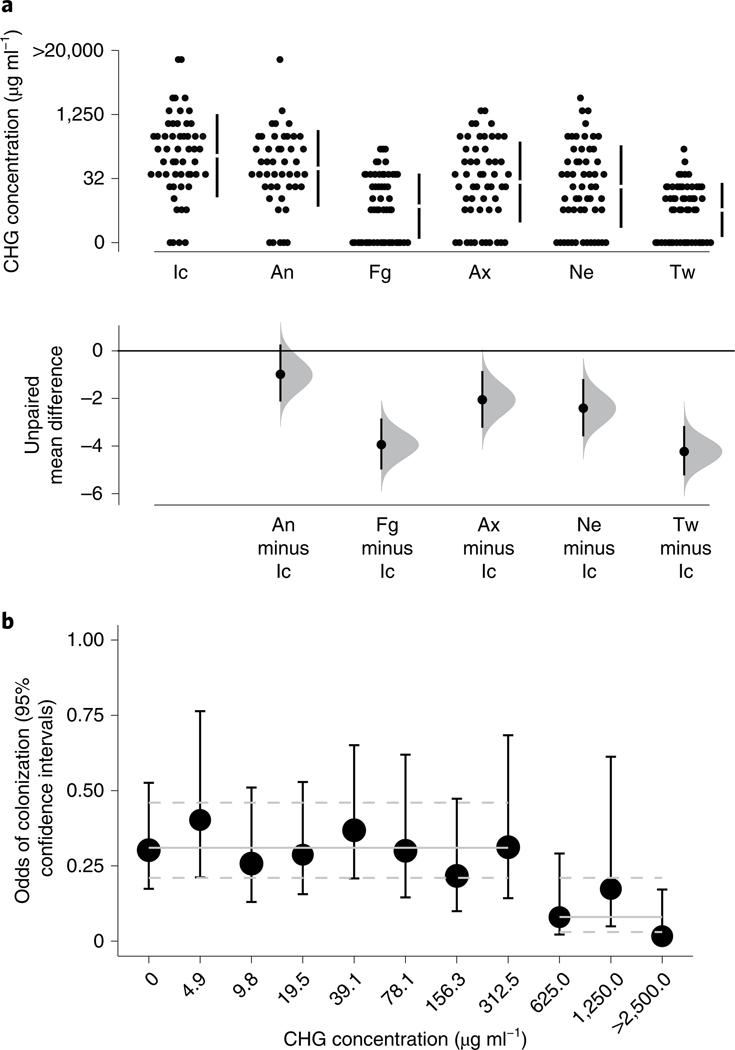
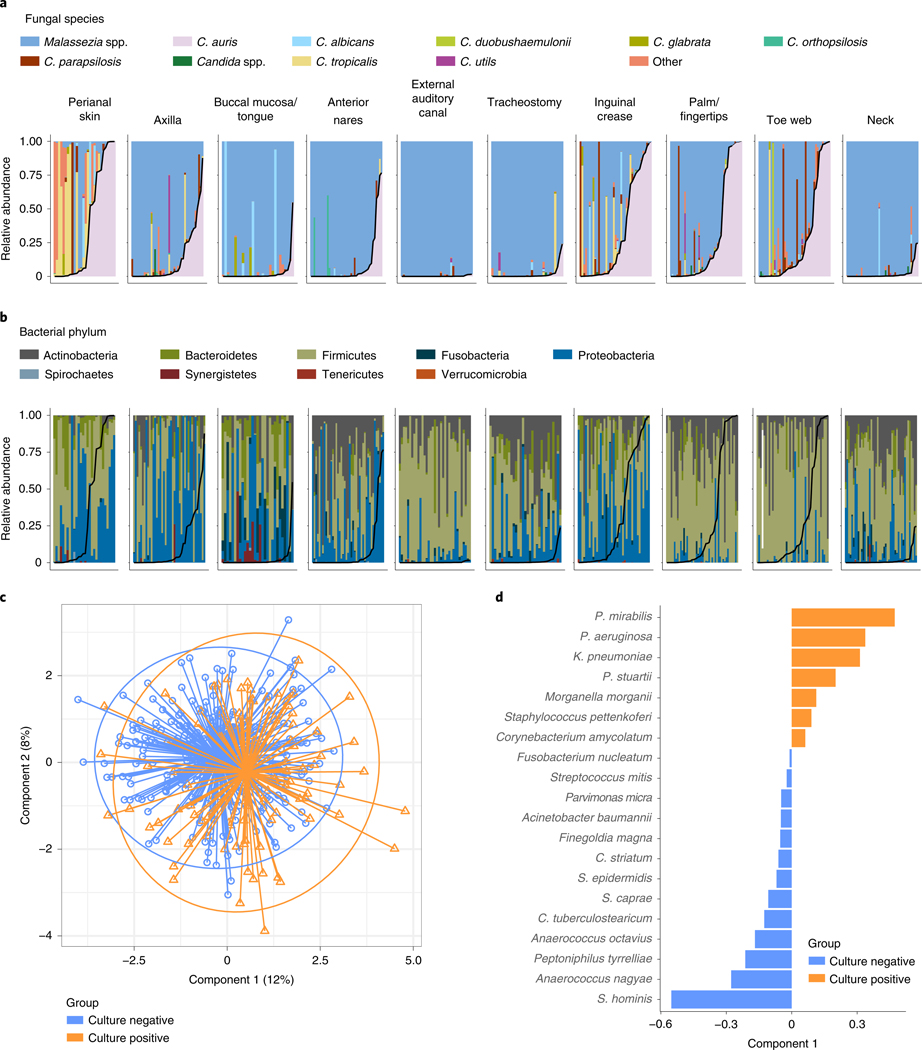
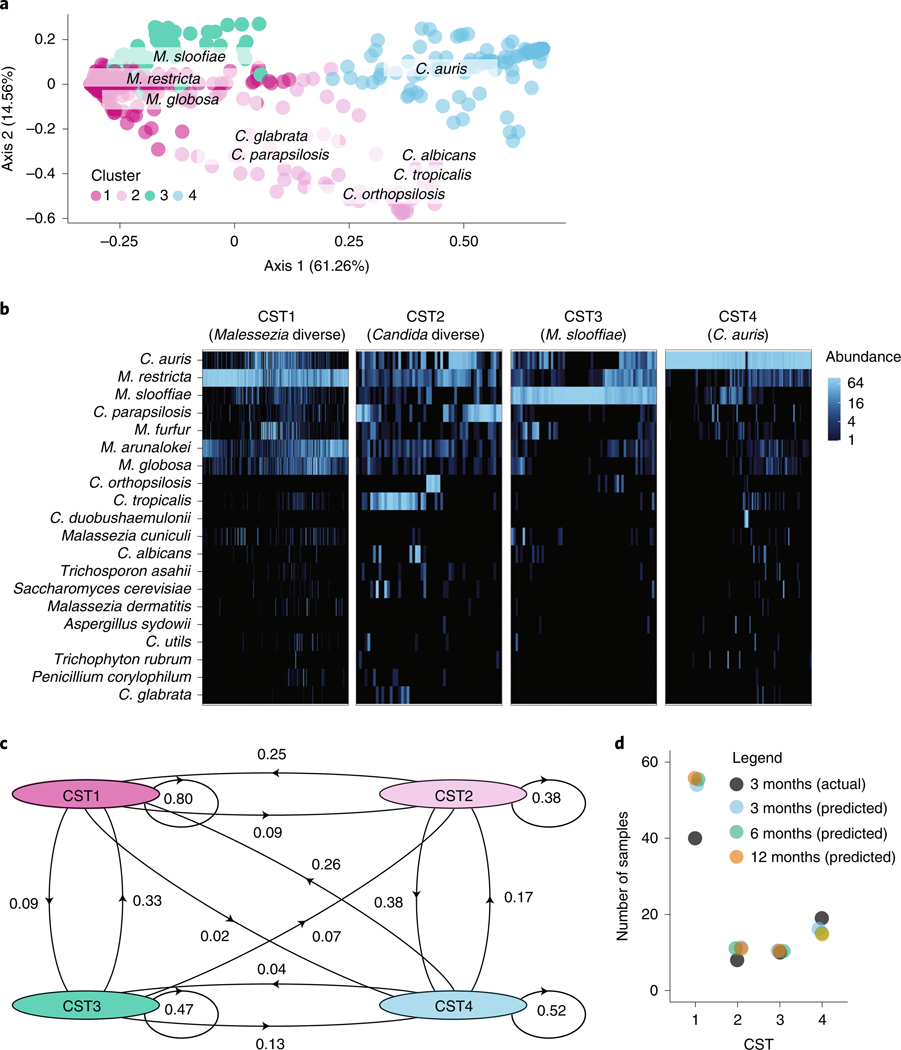
Candida auris is a fungal pathogen of high concern due to its ability to cause healthcare-associated infections and outbreaks, its resistance to antimicrobials and disinfectants and its persistence on human skin and in the inanimate environment. To inform surveillance and future mitigation strategies, we defined the extent of skin colonization and explored the microbiome associated with C. auris colonization. We collected swab specimens and clinical data at three times points between January and April 2019 from 57 residents (up to ten body sites each) of a ventilator-capable skilled nursing facility with endemic C. auris and routine chlorhexidine gluconate (CHG) bathing. Integrating microbial-genomic and epidemiologic data revealed occult C. auris colonization of multiple body sites not targeted commonly for screening. High concentrations of CHG were associated with suppression of C. auris growth but not with deleterious perturbation of commensal microbes. Modeling human mycobiome dynamics provided insight into underlying alterations to the skin fungal community as a possible modifiable risk factor for acquisition and persistence of C. auris. Failure to detect the extensive, disparate niches of C. auris colonization may reduce the effectiveness of infection-prevention measures that target colonized residents, highlighting the importance of universal strategies to reduce C. auris transmission.
© 2021. This is a U.S. government work and not under copyright protection in the U.S.; foreign copyright protection may apply.
Figures
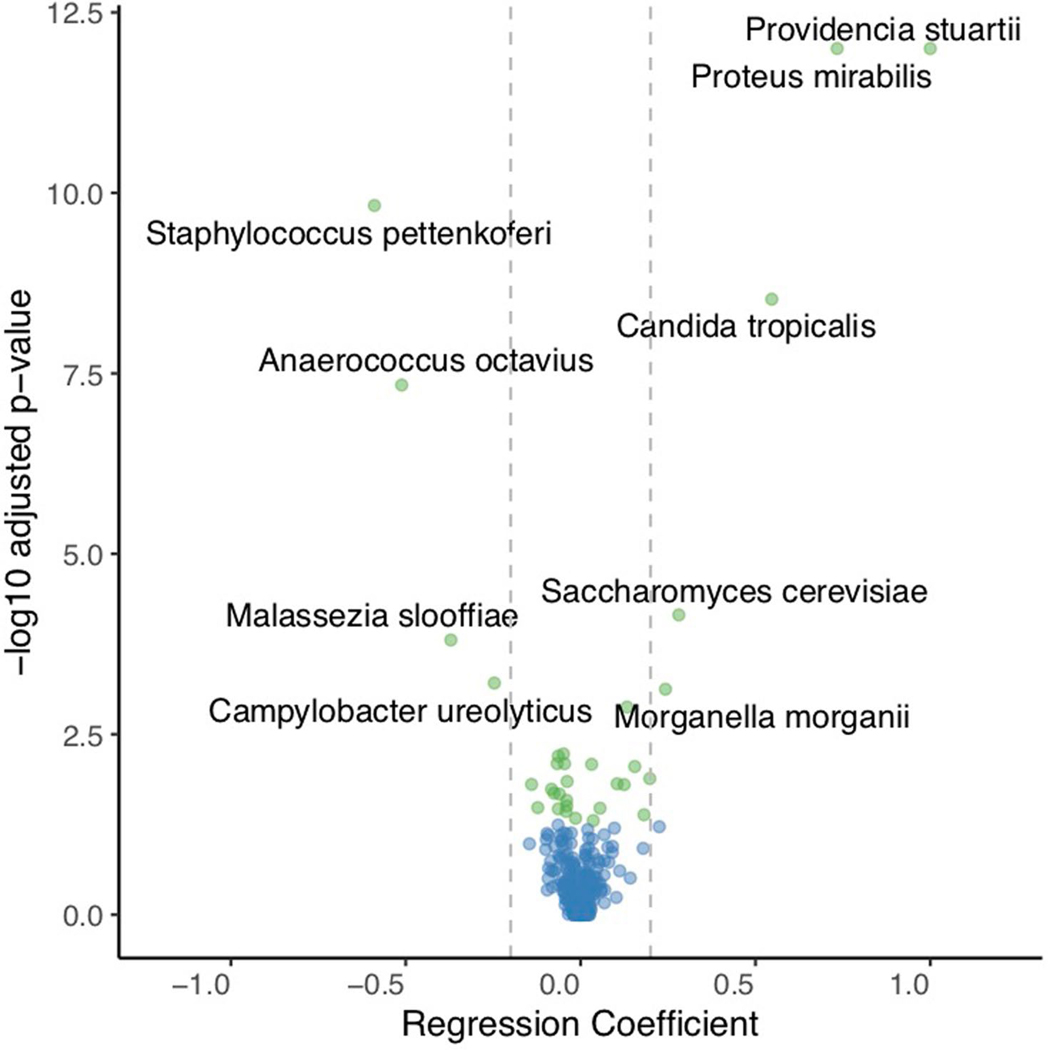
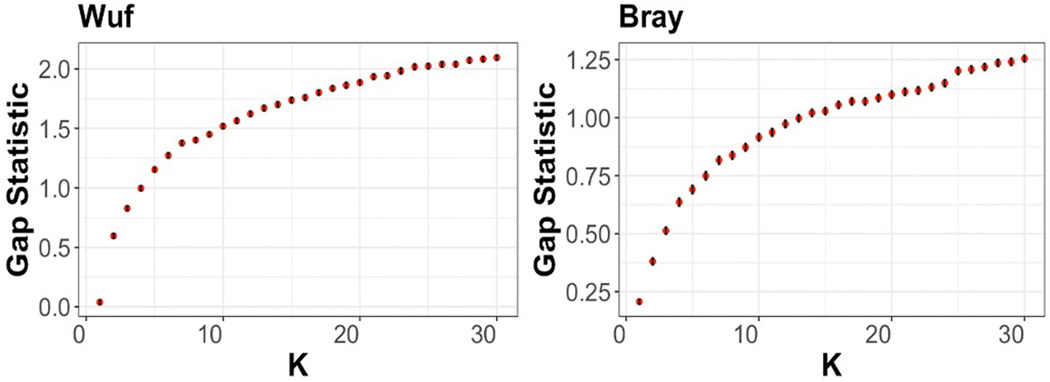
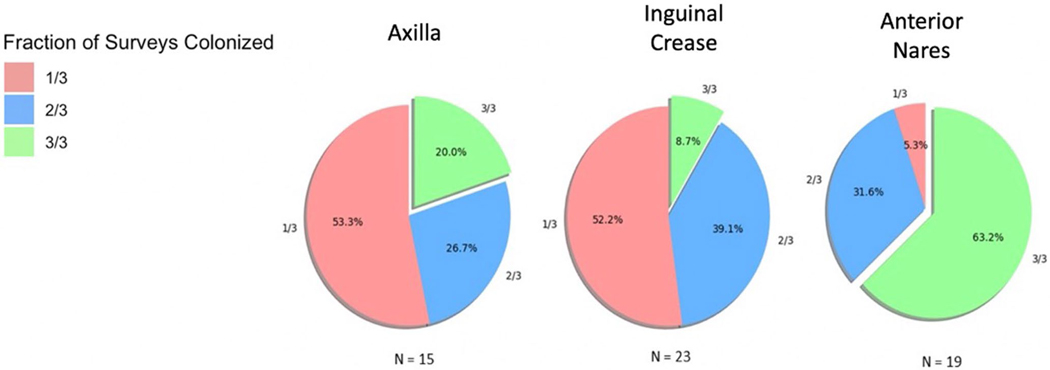
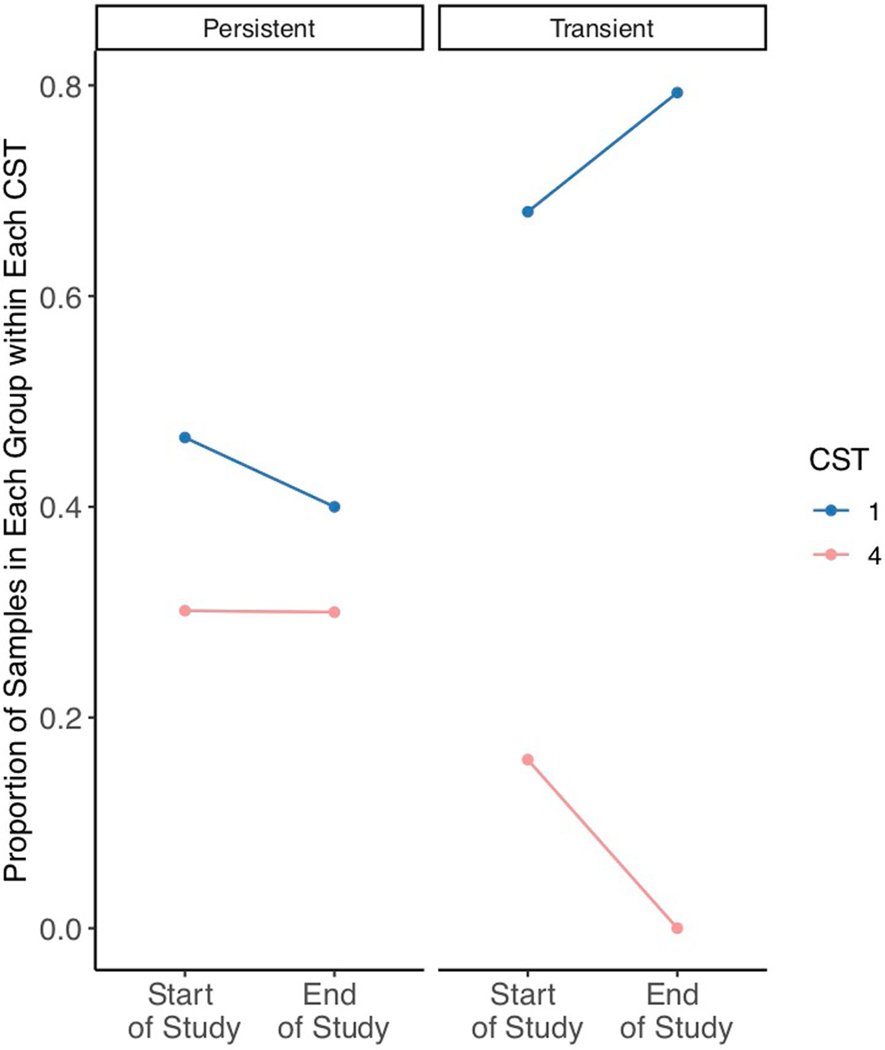
References
-
- Vallabhaneni S et al. Investigation of the first seven reported cases of Candida auris, a globally emerging invasive, multidrug-resistant fungus—United States, May 2013–August 2016. Am. J. Transplant. 17, 296–299 (2017). - PubMed
-
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019 (US Department of Health and Human Services, CDC, 2019). .
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
