

Molecular characterization of both transesterification reactions of the group II intron circularization pathway
- PMID: 34157113
- PMCID: PMC8266578
- DOI: 10.1093/nar/gkab537
Molecular characterization of both transesterification reactions of the group II intron circularization pathway
Abstract
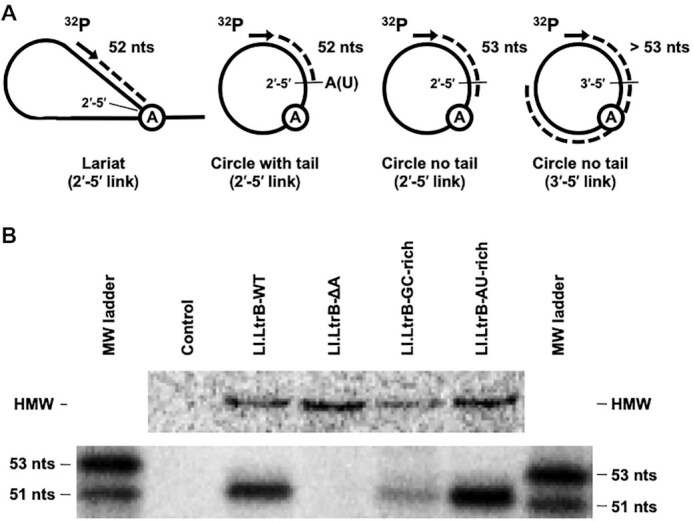
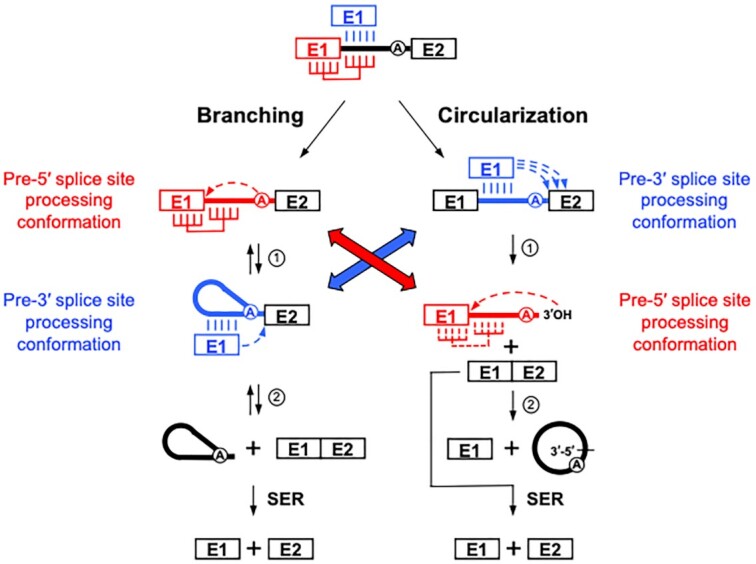
Group II introns can self-splice from RNA transcripts through branching, hydrolysis and circularization, being released as lariats, linear introns and circles, respectively. In contrast to branching, the circularization pathway is mostly based on assumptions and has been largely overlooked. Here, we address the molecular details of both transesterification reactions of the group II intron circularization pathway in vivo. We show that free E1 is recruited by the intron through base pairing interactions and that released intron circles can generate free E1 by the spliced exon reopening reaction. The first transesterification reaction was found to be induced inaccurately by the 3'OH of the terminal residue of free E1 at the 3' splice site, producing circularization intermediates with heterogeneous 3' ends. Nevertheless, specific terminal 3'OH, selected by a molecular ruler, was shown to precisely attack the 5' splice site and release intron circles with 3'-5' rather than 2'-5' bonds at their circularization junction. Our work supports a circularization model where the recruitment of free E1 and/or displacement of cis-E1 induce a conformational change of the intron active site from the pre-5' to the pre-3' splice site processing conformation, suggesting how circularization might initiate at the 3' instead of the 5' splice site.
© The Author(s) 2021. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures





Similar articles
-
Circularization pathway of a bacterial group II intron.Nucleic Acids Res. 2016 Feb 29;44(4):1845-53. doi: 10.1093/nar/gkv1381. Epub 2015 Dec 15. Nucleic Acids Res. 2016. PMID: 26673697 Free PMC article.
-
The Ll.LtrB intron from Lactococcus lactis excises as circles in vivo: insights into the group II intron circularization pathway.RNA. 2015 Jul;21(7):1286-93. doi: 10.1261/rna.046367.114. Epub 2015 May 8. RNA. 2015. PMID: 25956521 Free PMC article.
-
The ability to form full-length intron RNA circles is a general property of nuclear group I introns.RNA. 2003 Dec;9(12):1464-75. doi: 10.1261/rna.5290903. RNA. 2003. PMID: 14624003 Free PMC article.
-
Group II introns: structure and catalytic versatility of large natural ribozymes.Crit Rev Biochem Mol Biol. 2003;38(3):249-303. doi: 10.1080/713609236. Crit Rev Biochem Mol Biol. 2003. PMID: 12870716 Review.
-
Structure and activities of group II introns.Annu Rev Biochem. 1995;64:435-61. doi: 10.1146/annurev.bi.64.070195.002251. Annu Rev Biochem. 1995. PMID: 7574489 Review.
Cited by
-
Circular RNAs in the Origin of Developmental Lung Disease: Promising Diagnostic and Therapeutic Biomarkers.Biomolecules. 2023 Mar 15;13(3):533. doi: 10.3390/biom13030533. Biomolecules. 2023. PMID: 36979468 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
