

Overexpressing low-density lipoprotein receptor reduces tau-associated neurodegeneration in relation to apoE-linked mechanisms
- PMID: 34157306
- PMCID: PMC8349883
- DOI: 10.1016/j.neuron.2021.05.034
Overexpressing low-density lipoprotein receptor reduces tau-associated neurodegeneration in relation to apoE-linked mechanisms
Abstract
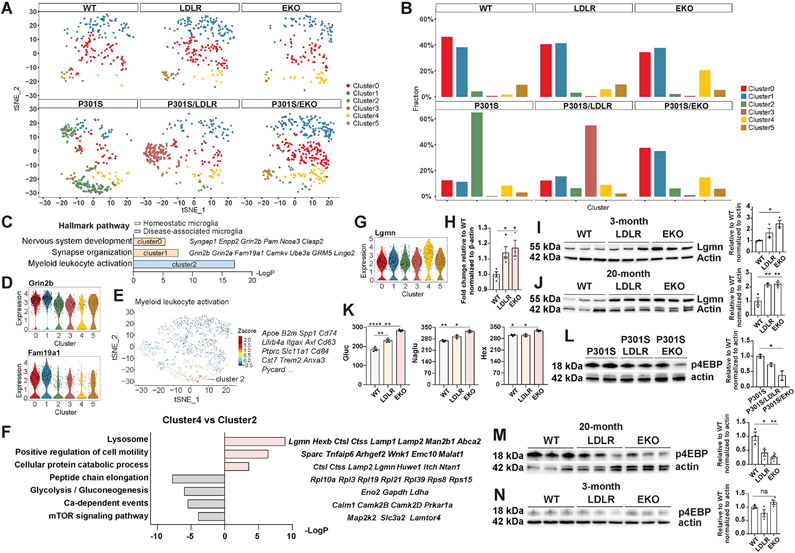
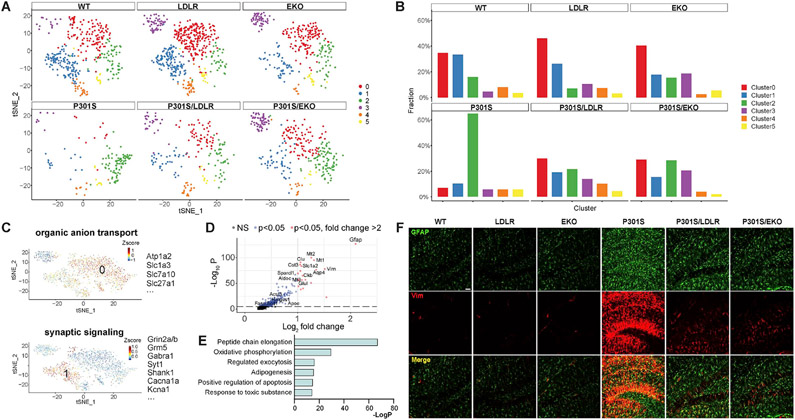
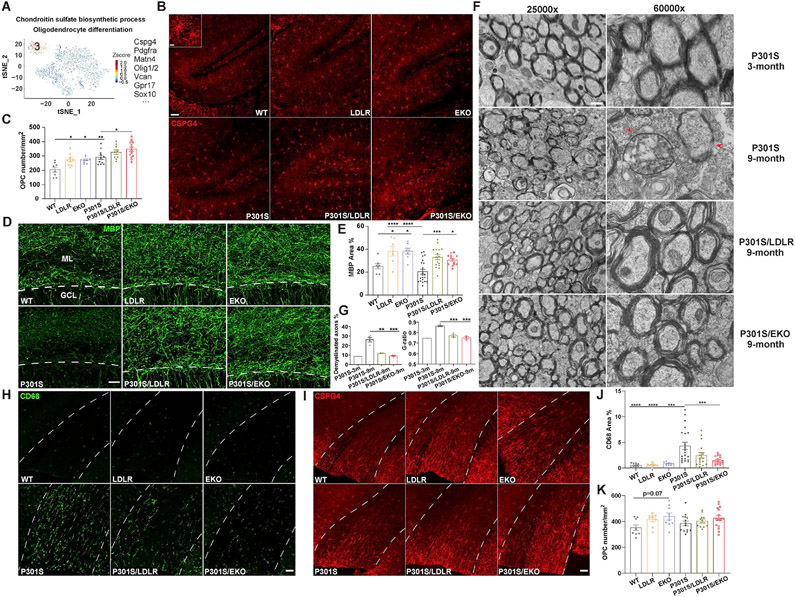
APOE is the strongest genetic risk factor for late-onset Alzheimer's disease. ApoE exacerbates tau-associated neurodegeneration by driving microglial activation. However, how apoE regulates microglial activation and whether targeting apoE is therapeutically beneficial in tauopathy is unclear. Here, we show that overexpressing an apoE metabolic receptor, LDLR (low-density lipoprotein receptor), in P301S tauopathy mice markedly reduces brain apoE and ameliorates tau pathology and neurodegeneration. LDLR overexpression (OX) in microglia cell-autonomously downregulates microglial Apoe expression and is associated with suppressed microglial activation as in apoE-deficient microglia. ApoE deficiency and LDLR OX strongly drive microglial immunometabolism toward enhanced catabolism over anabolism, whereas LDLR-overexpressing microglia also uniquely upregulate specific ion channels and neurotransmitter receptors upon activation. ApoE-deficient and LDLR-overexpressing mice harbor enlarged pools of oligodendrocyte progenitor cells (OPCs) and show greater preservation of myelin integrity under neurodegenerative conditions. They also show less reactive astrocyte activation in the setting of tauopathy.
Keywords: ApoE; LDLR; OPC; metabolism; microglia; myelin; tau.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests D.M.H. and C.I. are listed as inventors on a patent licensed by Washington University to C2N Diagnostics on the therapeutic use of anti-tau antibodies. D.M.H. co-founded and is on the scientific advisory board of C2N Diagnostics. C2N Diagnostics has licensed certain anti-tau antibodies to AbbVie for therapeutic development. D.M.H. is on the scientific advisory board of Denali and consults for Genentech, Merck, and Cajal Neurosciences.
Figures
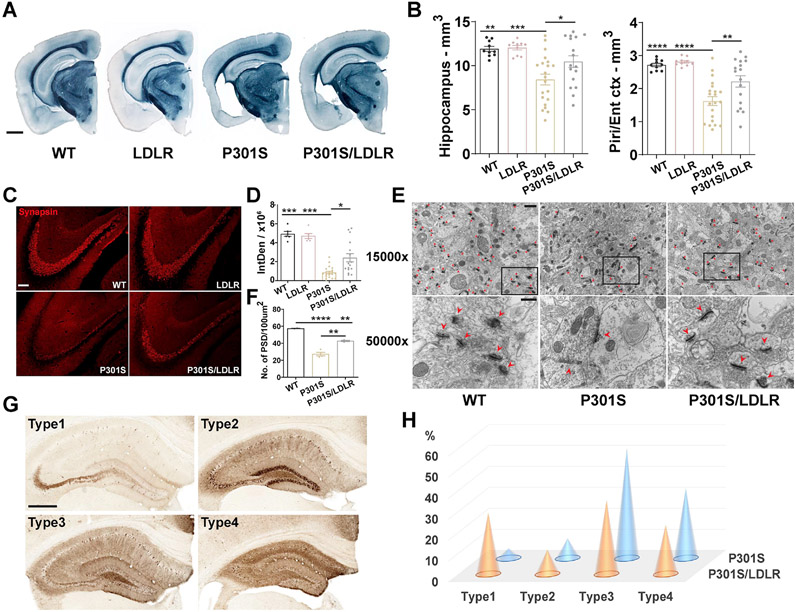
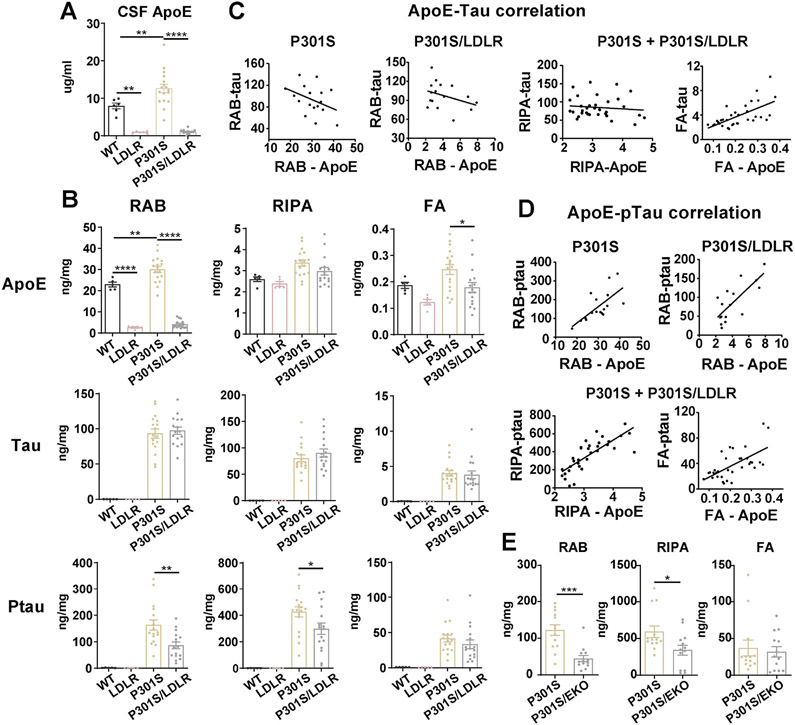
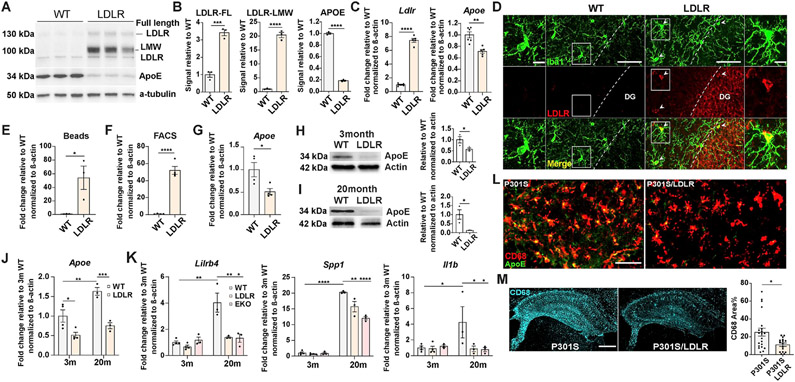
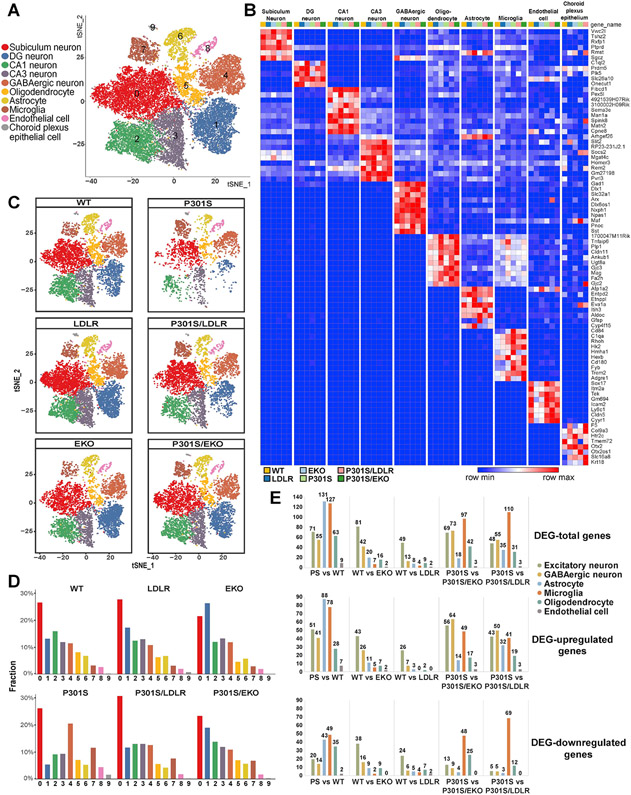
Comment in
-
ApoE and immunity in Alzheimer's disease and related tauopathies: Low-density lipoprotein receptor to the rescue.Neuron. 2021 Aug 4;109(15):2363-2365. doi: 10.1016/j.neuron.2021.07.013. Neuron. 2021. PMID: 34352209 Free PMC article.
References
-
- Borchelt DR, Davis J, Fischer M, Lee MK, Slunt HH, Ratovitsky T, Regard J, Copeland NG, Jenkins NA, Sisodia SS, and Price DL (1996). A vector for expressing foreign genes in the brains and hearts of transgenic mice. Genetic Analysis: Biomolecular Engineering 13, 159–163. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
