Sperm mosaicism: implications for genomic diversity and disease
- PMID: 34158173
- PMCID: PMC9484299
- DOI: 10.1016/j.tig.2021.05.007
Sperm mosaicism: implications for genomic diversity and disease
Abstract
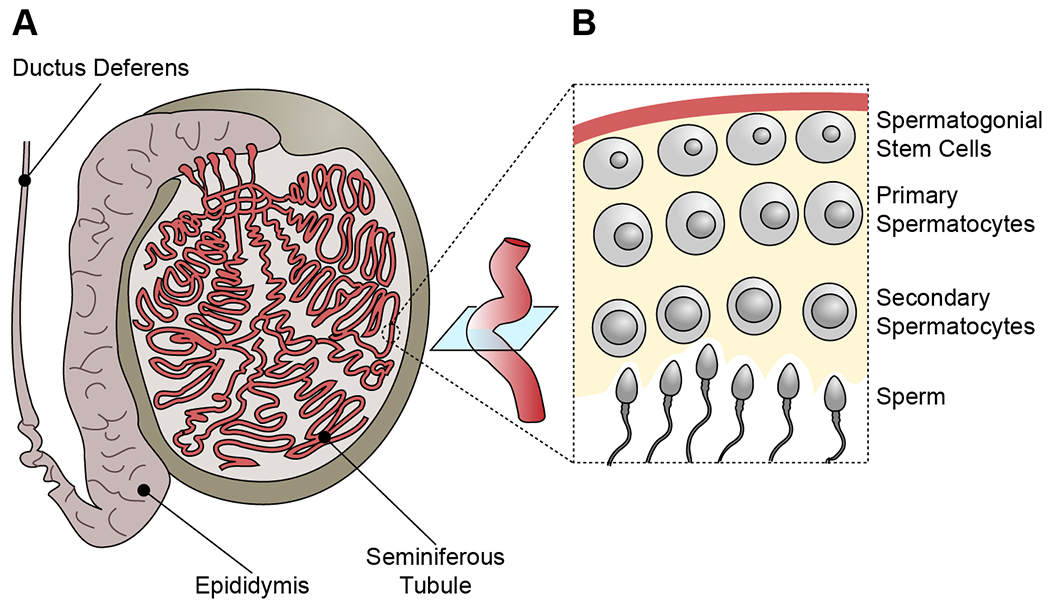
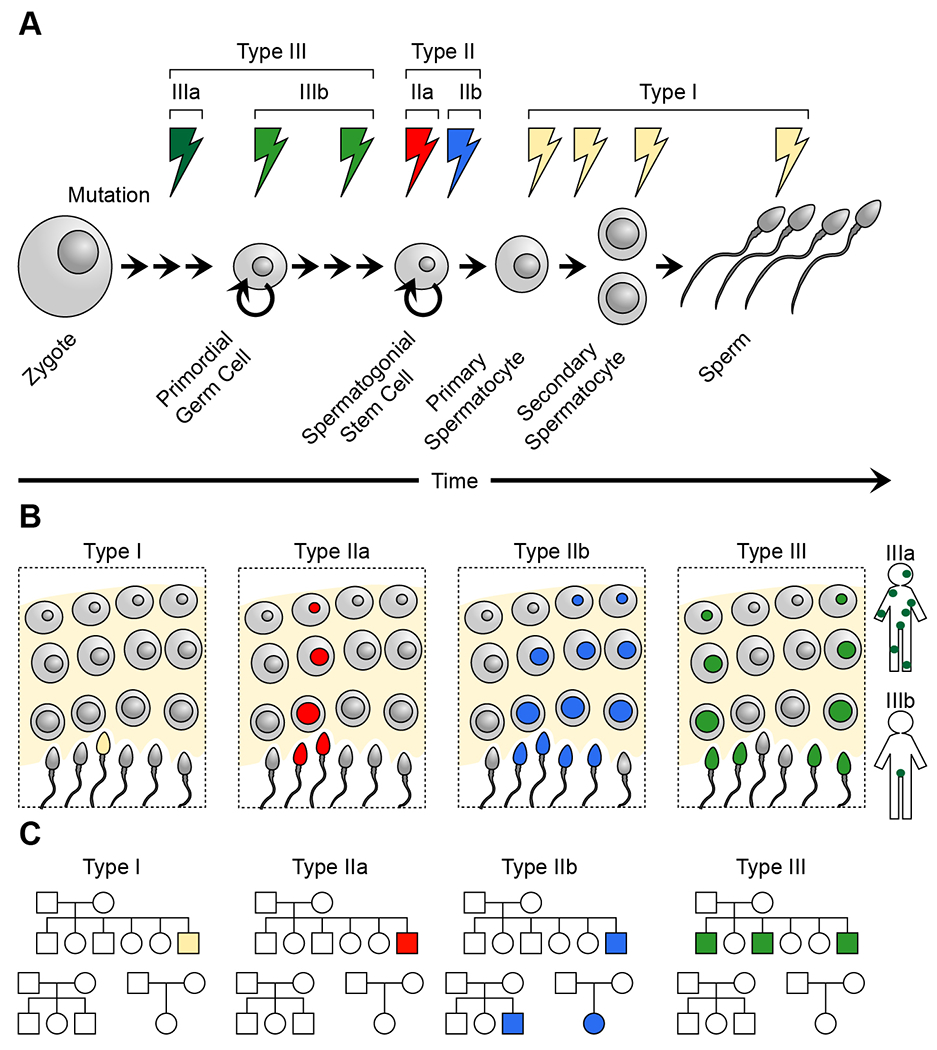
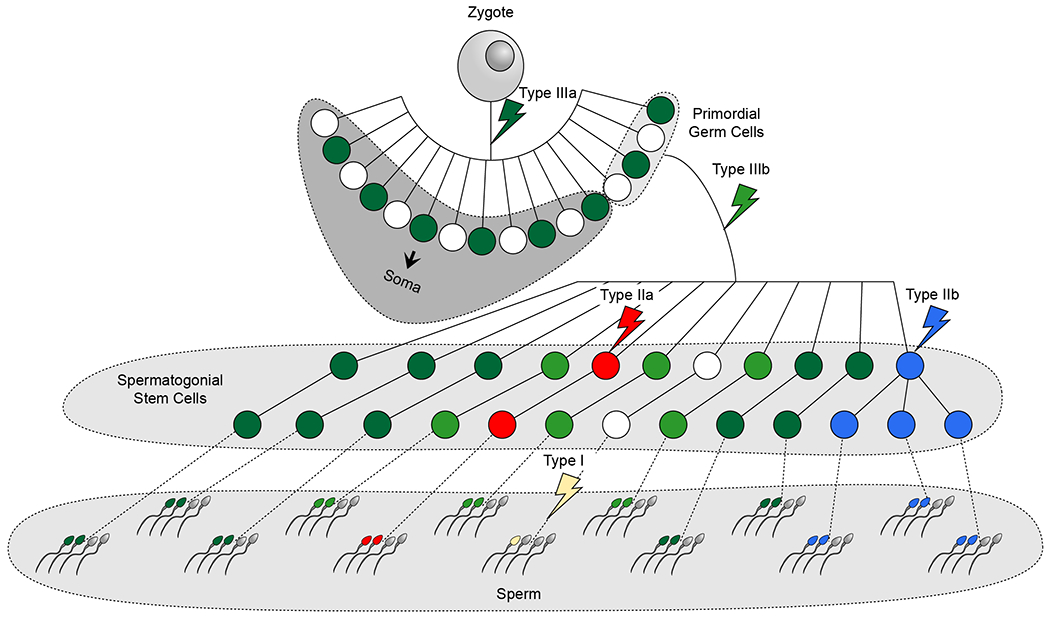
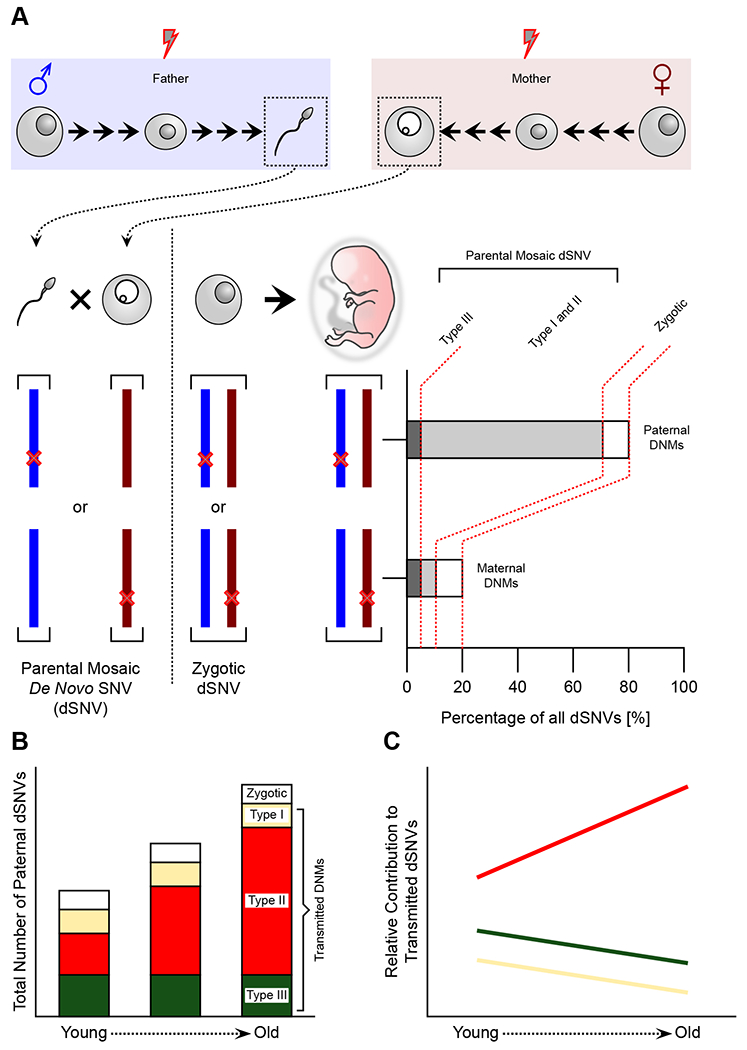
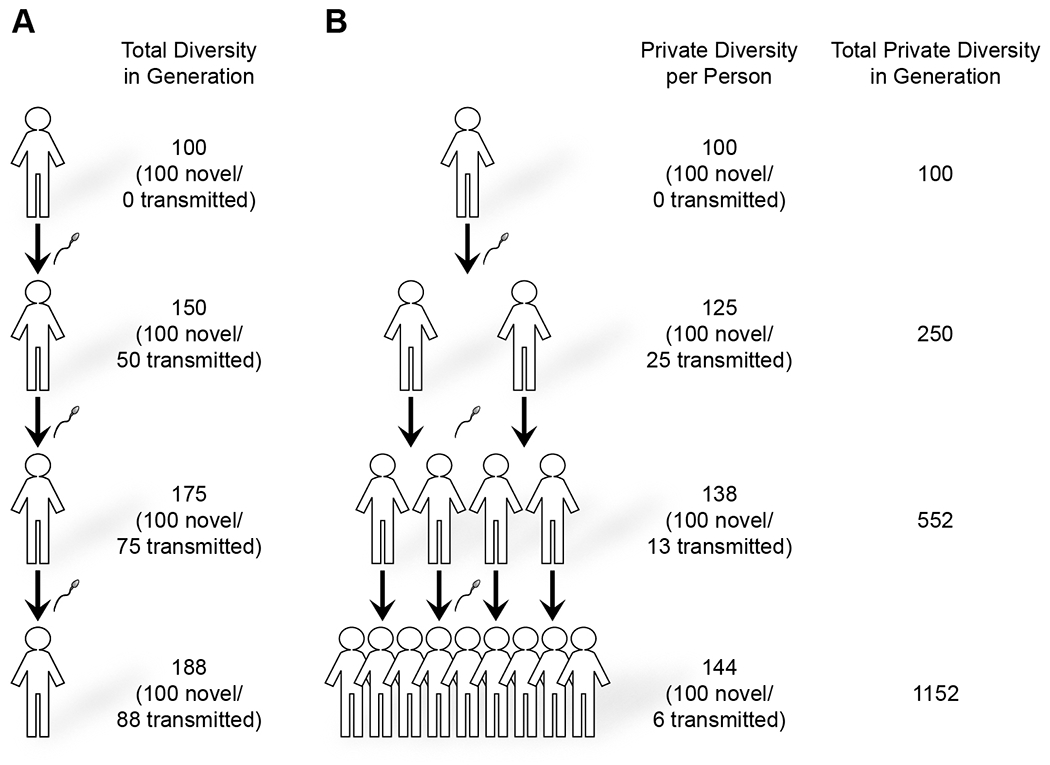
While sperm mosaicism has few consequences for men, the offspring and future generations are unwitting recipients of gonadal cell mutations, often yielding severe disease. Recent studies, fueled by emergent technologies, show that sperm mosaicism is a common source of de novo mutations (DNMs) that underlie severe pediatric disease as well as human genetic diversity. Sperm mosaicism can be divided into three types: Type I arises during sperm meiosis and is non-age dependent; Type II arises in spermatogonia and increases as men age; and Type III arises during paternal embryogenesis, spreads throughout the body, and contributes stably to sperm throughout life. Where Types I and II confer little risk of recurrence, Type III may confer identifiable risk to future offspring. These mutations are likely to be the single largest contributor to human genetic diversity. New sequencing approaches may leverage this framework to evaluate and reduce disease risk for future generations.
Keywords: advanced paternal age; human variation; primordial germ cells; sperm mosaicism; spermatogonial stem cells.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests No interests are declared. M.W.B. and J.G.G. are inventors on a patent (PCT/US2018/024878, WO2018183525A1) filed by UC, San Diego that is titled 'Methods for assessing risk of or diagnosing genetic defects by identifying de novo mutations or somatic mosaic variants in sperm or somatic tissues'.
Figures