

Rapid evolution of bacterial mutualism in the plant rhizosphere
- PMID: 34158504
- PMCID: PMC8219802
- DOI: 10.1038/s41467-021-24005-y
Rapid evolution of bacterial mutualism in the plant rhizosphere
Abstract
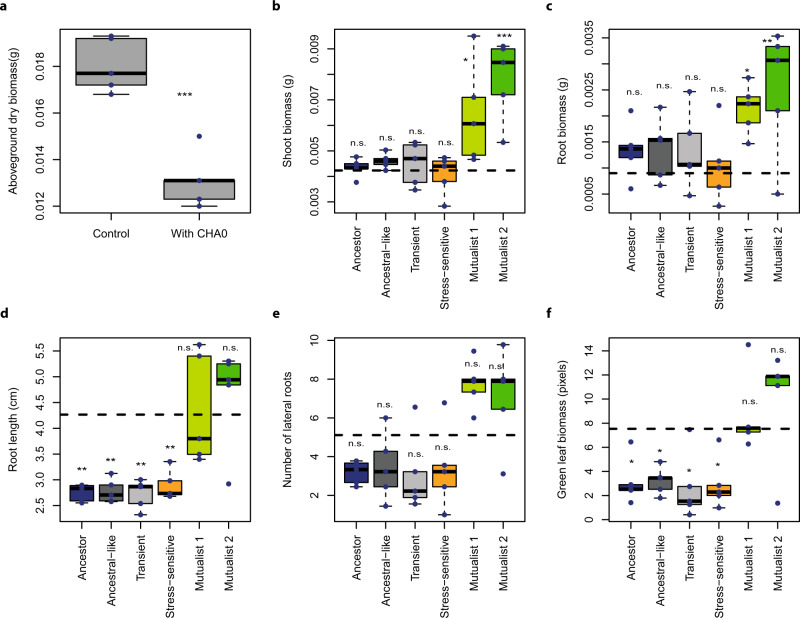
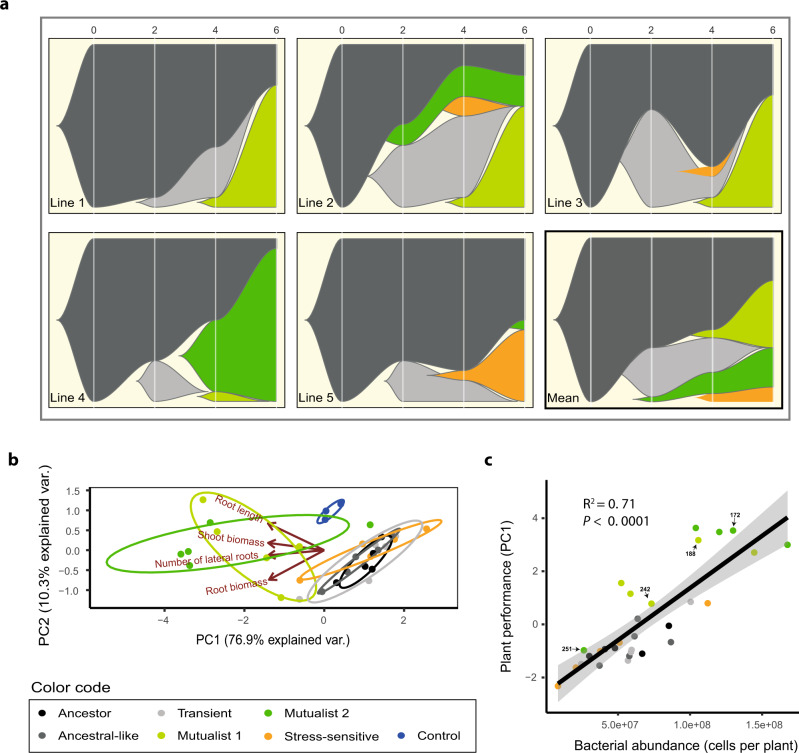
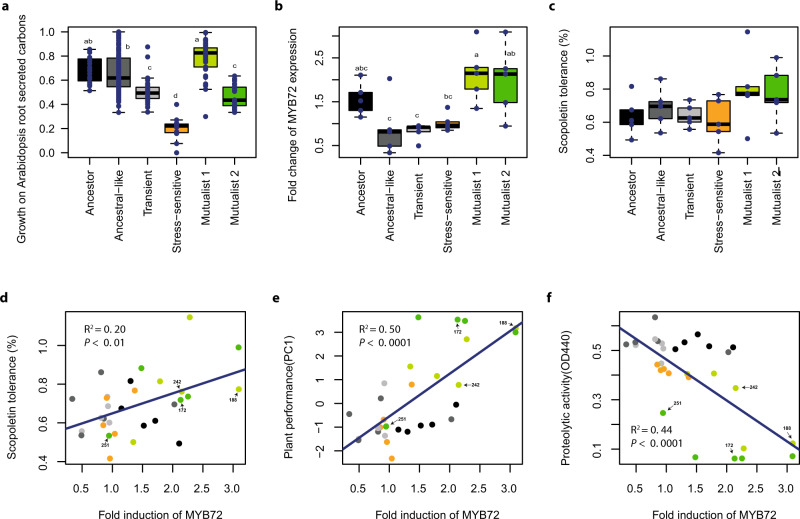
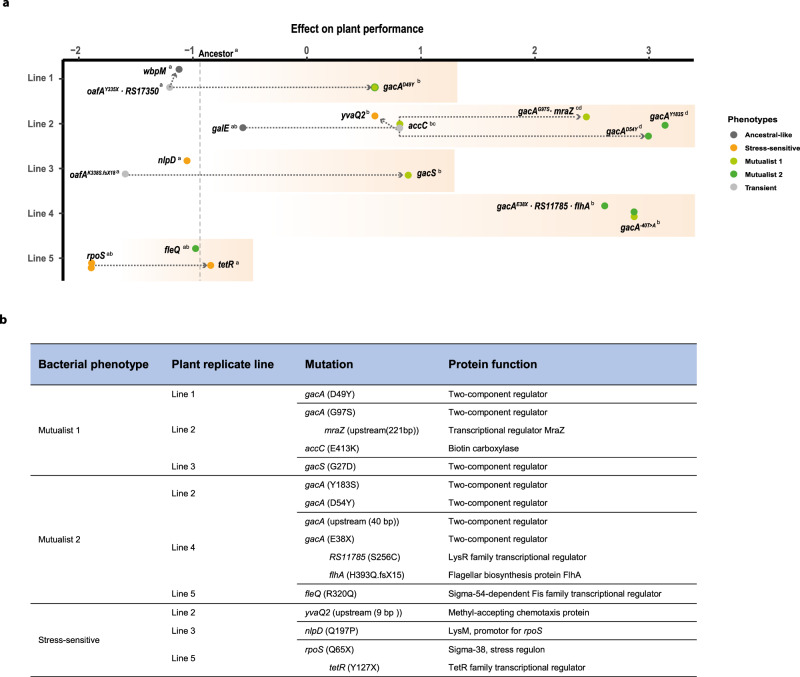
While beneficial plant-microbe interactions are common in nature, direct evidence for the evolution of bacterial mutualism is scarce. Here we use experimental evolution to causally show that initially plant-antagonistic Pseudomonas protegens bacteria evolve into mutualists in the rhizosphere of Arabidopsis thaliana within six plant growth cycles (6 months). This evolutionary transition is accompanied with increased mutualist fitness via two mechanisms: (i) improved competitiveness for root exudates and (ii) enhanced tolerance to the plant-secreted antimicrobial scopoletin whose production is regulated by transcription factor MYB72. Crucially, these mutualistic adaptations are coupled with reduced phytotoxicity, enhanced transcription of MYB72 in roots, and a positive effect on plant growth. Genetically, mutualism is associated with diverse mutations in the GacS/GacA two-component regulator system, which confers high fitness benefits only in the presence of plants. Together, our results show that rhizosphere bacteria can rapidly evolve along the parasitism-mutualism continuum at an agriculturally relevant evolutionary timescale.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
β-Glucosidase BGLU42 is a MYB72-dependent key regulator of rhizobacteria-induced systemic resistance and modulates iron deficiency responses in Arabidopsis roots.New Phytol. 2014 Oct;204(2):368-79. doi: 10.1111/nph.12980. Epub 2014 Aug 19. New Phytol. 2014. PMID: 25138267
-
Experimental-Evolution-Driven Identification of Arabidopsis Rhizosphere Competence Genes in Pseudomonas protegens.mBio. 2021 Jun 29;12(3):e0092721. doi: 10.1128/mBio.00927-21. Epub 2021 Jun 8. mBio. 2021. PMID: 34101491 Free PMC article.
-
MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health.Proc Natl Acad Sci U S A. 2018 May 29;115(22):E5213-E5222. doi: 10.1073/pnas.1722335115. Epub 2018 Apr 23. Proc Natl Acad Sci U S A. 2018. PMID: 29686086 Free PMC article.
-
Dissecting endophytic lifestyle along the parasitism/mutualism continuum in Arabidopsis.Curr Opin Microbiol. 2016 Aug;32:103-112. doi: 10.1016/j.mib.2016.05.008. Epub 2016 Jun 6. Curr Opin Microbiol. 2016. PMID: 27280851 Review.
-
Housing helpful invaders: the evolutionary and molecular architecture underlying plant root-mutualist microbe interactions.J Exp Bot. 2015 Apr;66(8):2177-86. doi: 10.1093/jxb/erv038. Epub 2015 Mar 5. J Exp Bot. 2015. PMID: 25743160 Free PMC article. Review.
Cited by
-
Community standards and future opportunities for synthetic communities in plant-microbiota research.Nat Microbiol. 2024 Nov;9(11):2774-2784. doi: 10.1038/s41564-024-01833-4. Epub 2024 Oct 30. Nat Microbiol. 2024. PMID: 39478084 Review.
-
Effect ofBacillus velezensis JC-K3 on Endophytic Bacterial and Fungal Diversity in Wheat Under Salt Stress.Front Microbiol. 2021 Dec 20;12:802054. doi: 10.3389/fmicb.2021.802054. eCollection 2021. Front Microbiol. 2021. PMID: 34987493 Free PMC article.
-
Phenotypic and genomic signatures of interspecies cooperation and conflict in naturally occurring isolates of a model plant symbiont.Proc Biol Sci. 2022 Jul 13;289(1978):20220477. doi: 10.1098/rspb.2022.0477. Epub 2022 Jul 13. Proc Biol Sci. 2022. PMID: 35858063 Free PMC article.
-
Co-evolution within the plant holobiont drives host performance.EMBO Rep. 2023 Sep 6;24(9):e57455. doi: 10.15252/embr.202357455. Epub 2023 Jul 20. EMBO Rep. 2023. PMID: 37471099 Free PMC article. Review.
-
Characterization of the Cultivable Endophytic Bacterial Community of Seeds and Sprouts of Cannabis sativa L. and Perspectives for the Application as Biostimulants.Microorganisms. 2022 Aug 29;10(9):1742. doi: 10.3390/microorganisms10091742. Microorganisms. 2022. PMID: 36144344 Free PMC article.
References
-
- Kiers ET, Denison RF. Sanctions, cooperation, and the stability of plant-rhizosphere mutualisms. Annu. Rev. Ecol. Evol. Syst. 2008;39:215–236. doi: 10.1146/annurev.ecolsys.39.110707.173423. - DOI
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
