

Introgression of the sesquiterpene biosynthesis from Solanum habrochaites to cultivated tomato offers insights into trichome morphology and arthropod resistance
- PMID: 34160697
- PMCID: PMC8222033
- DOI: 10.1007/s00425-021-03651-y
Introgression of the sesquiterpene biosynthesis from Solanum habrochaites to cultivated tomato offers insights into trichome morphology and arthropod resistance
Abstract
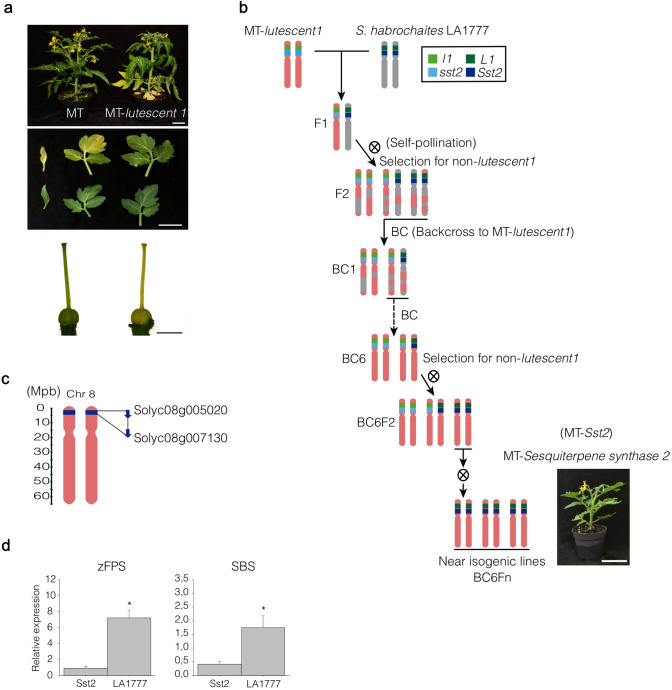
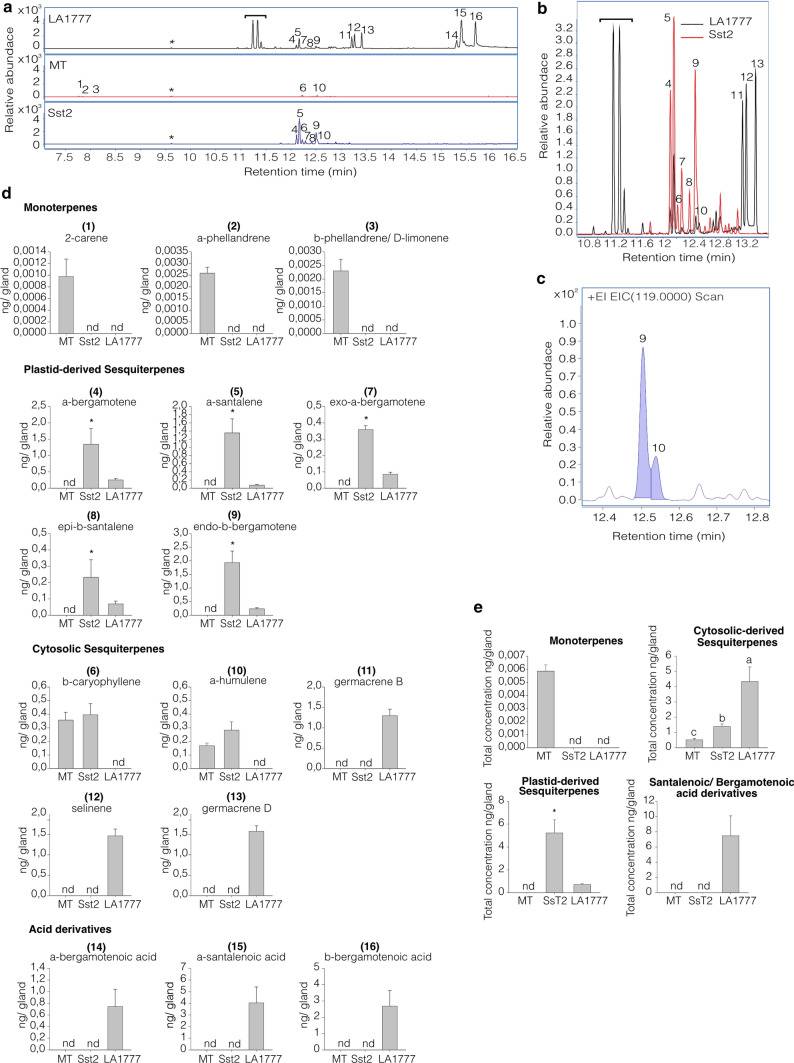
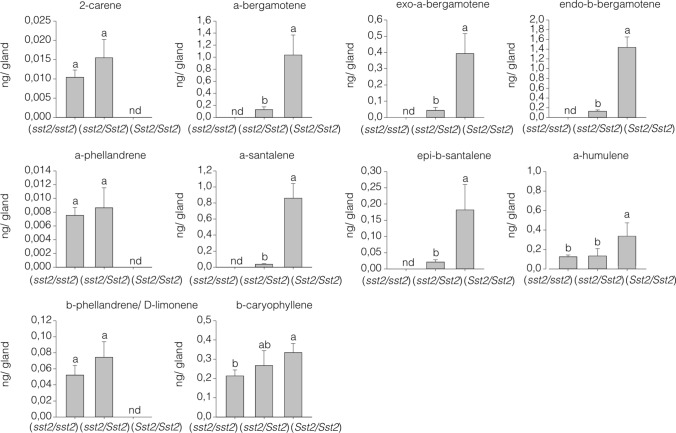
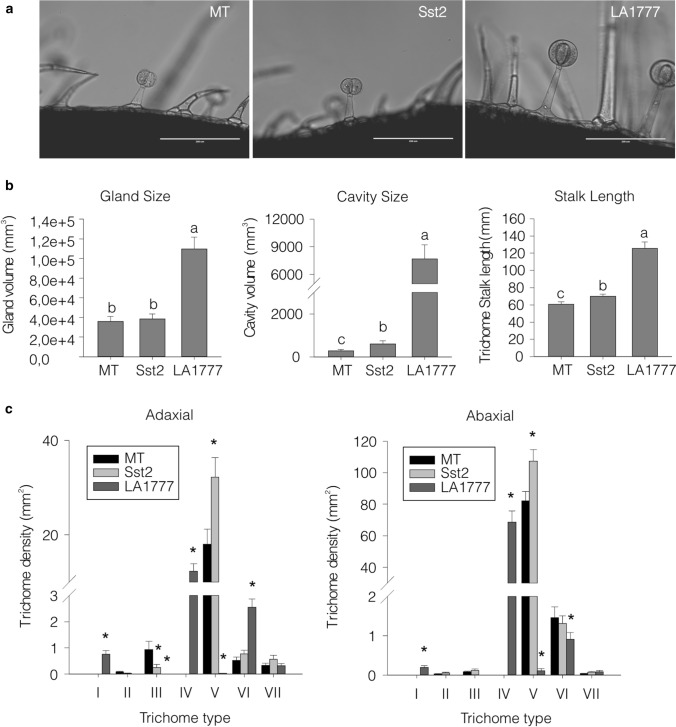
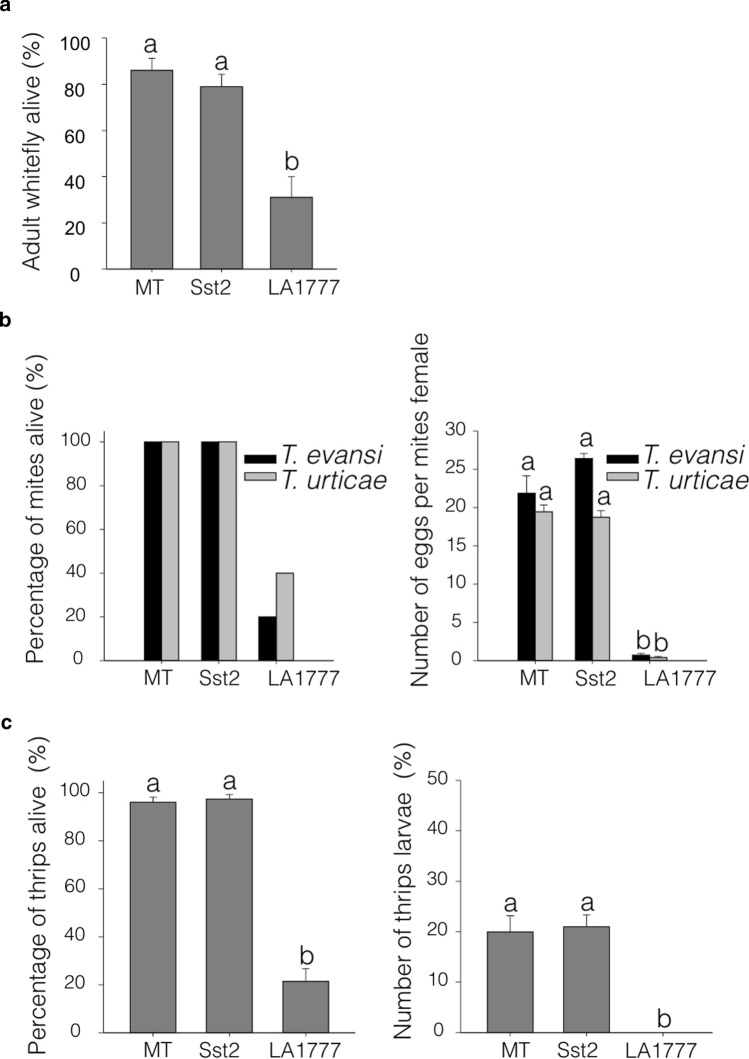
Cultivated tomatoes harboring the plastid-derived sesquiterpenes from S. habrochaites have altered type-VI trichome morphology and unveil additional genetic components necessary for piercing-sucking pest resistance. Arthropod resistance in the tomato wild relative Solanum habrochaites LA1777 is linked to specific sesquiterpene biosynthesis. The Sesquiterpene synthase 2 (SsT2) gene cluster on LA1777 chromosome 8 controls plastid-derived sesquiterpene synthesis. The main genes at SsT2 are Z-prenyltransferase (zFPS) and Santalene and Bergamotene Synthase (SBS), which produce α-santalene, β-bergamotene, and α-bergamotene in LA1777 round-shaped type-VI glandular trichomes. Cultivated tomatoes have mushroom-shaped type-VI trichomes with much smaller glands that contain low levels of monoterpenes and cytosolic-derived sesquiterpenes, not presenting the same pest resistance as in LA1777. We successfully transferred zFPS and SBS from LA1777 to cultivated tomato (cv. Micro-Tom, MT) by a backcrossing approach. The trichomes of the MT-Sst2 introgressed line produced high levels of the plastid-derived sesquiterpenes. The type-VI trichome internal storage-cavity size increased in MT-Sst2, probably as an effect of the increased amount of sesquiterpenes, although it was not enough to mimic the round-shaped LA1777 trichomes. The presence of high amounts of plastid-derived sesquiterpenes was also not sufficient to confer resistance to various tomato piercing-sucking pests, indicating that the effect of the sesquiterpenes found in the wild S. habrochaites can be insect specific. Our results provide for a better understanding of the morphology of S. habrochaites type-VI trichomes and paves the way to obtain insect-resistant tomatoes.
Keywords: Bergamotene; Glandular trichome; Introgressed line; Piercing-sucking pest; Santalene; Terpenes; Tomato trichome.
Figures
References
MeSH terms
Substances
Grants and funding
- 2016/22323-4/Fundação de Amparo à Pesquisa do Estado de São Paulo
- 2018/05003-1/Fundação de Amparo à Pesquisa do Estado de São Paulo
- 88881.189015/2018-01/Coordenação de Aperfeiçoamento de Pessoal de Nível Superior
- 307893/2016-2/Conselho Nacional de Desenvolvimento Científico e Tecnológico
- 306518/2018-0/Conselho Nacional de Desenvolvimento Científico e Tecnológico
LinkOut - more resources
Full Text Sources
