

Monocarboxylate transporter antagonism reveals metabolic vulnerabilities of viral-driven lymphomas
- PMID: 34161263
- PMCID: PMC8237662
- DOI: 10.1073/pnas.2022495118
Monocarboxylate transporter antagonism reveals metabolic vulnerabilities of viral-driven lymphomas
Abstract
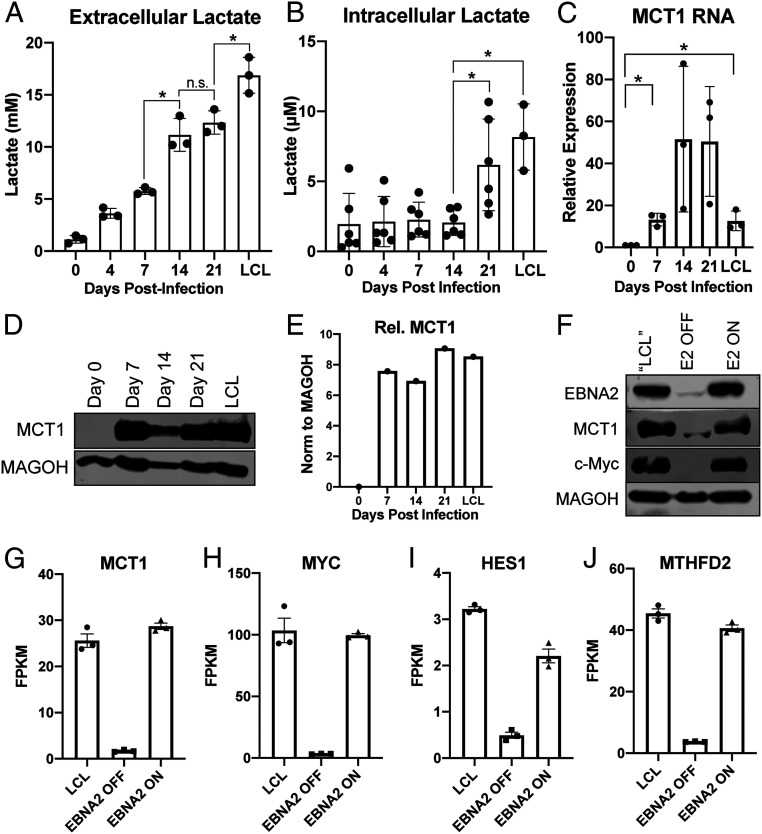
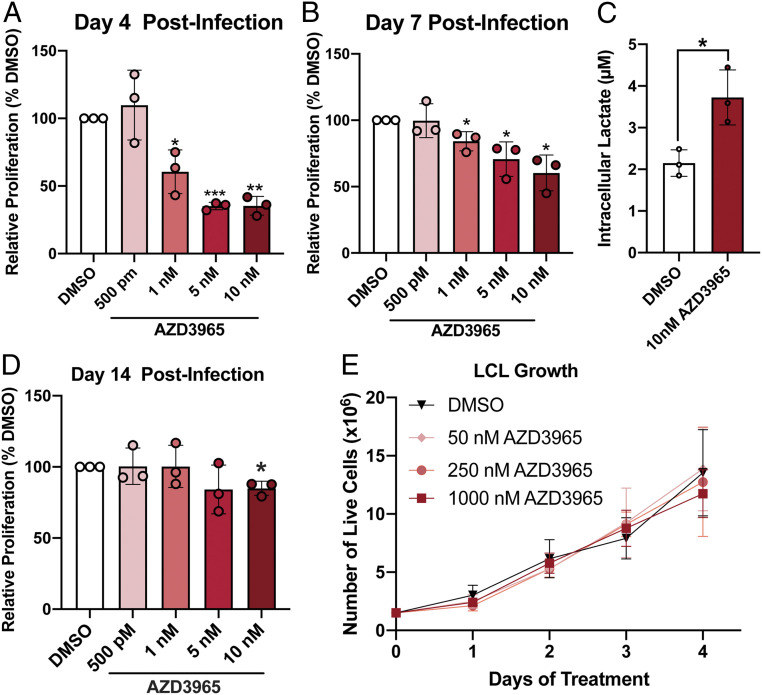
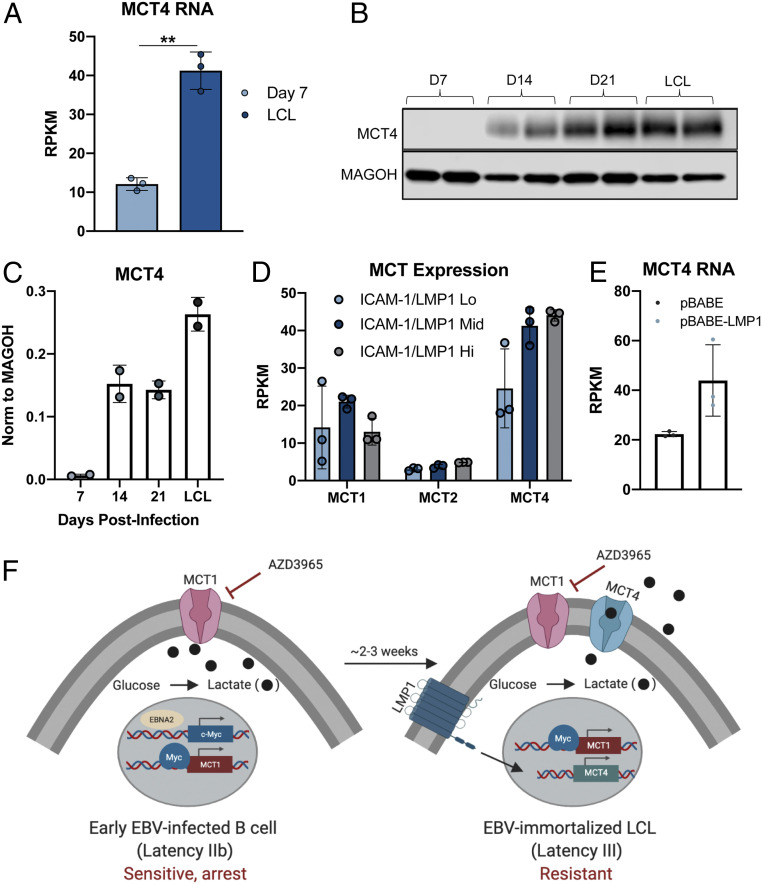
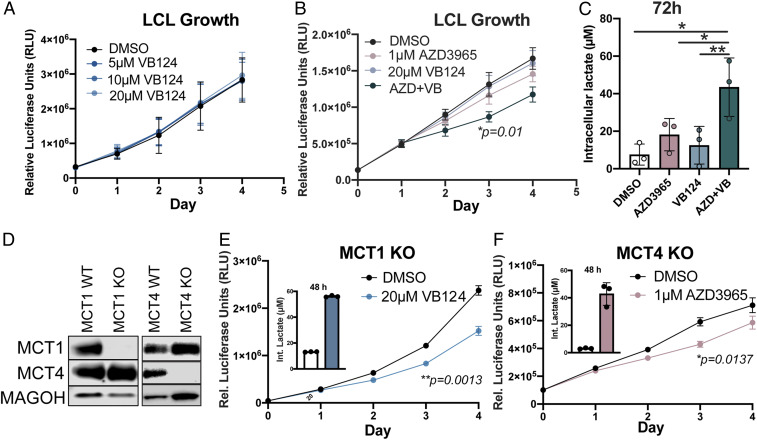
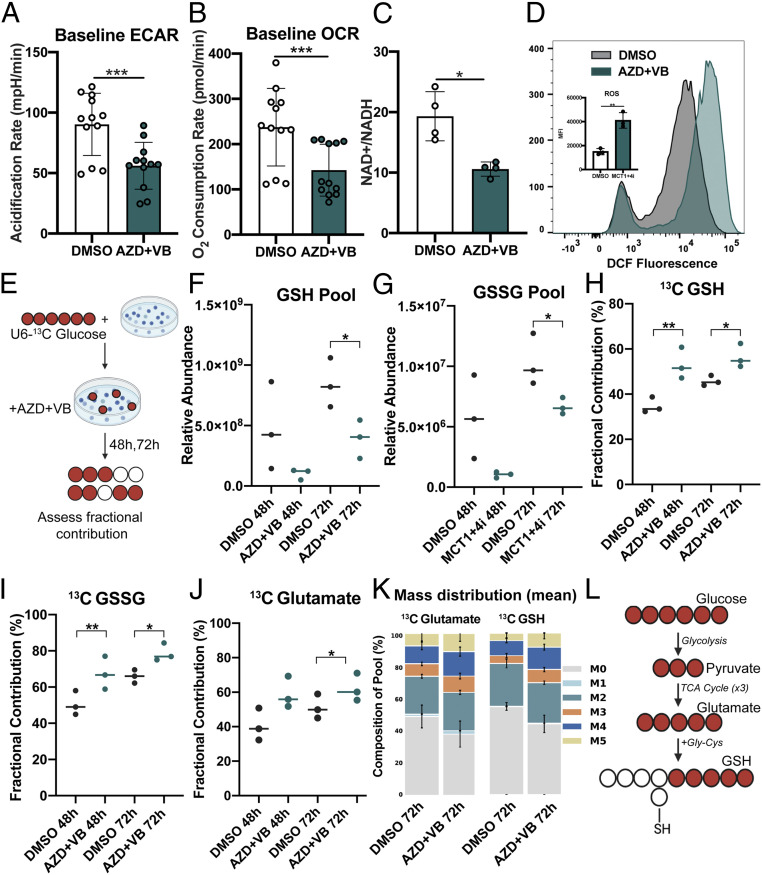
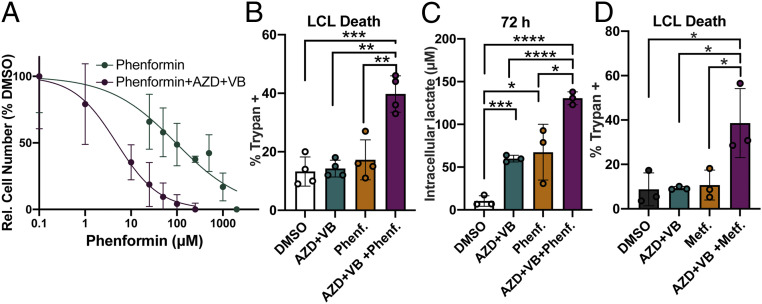
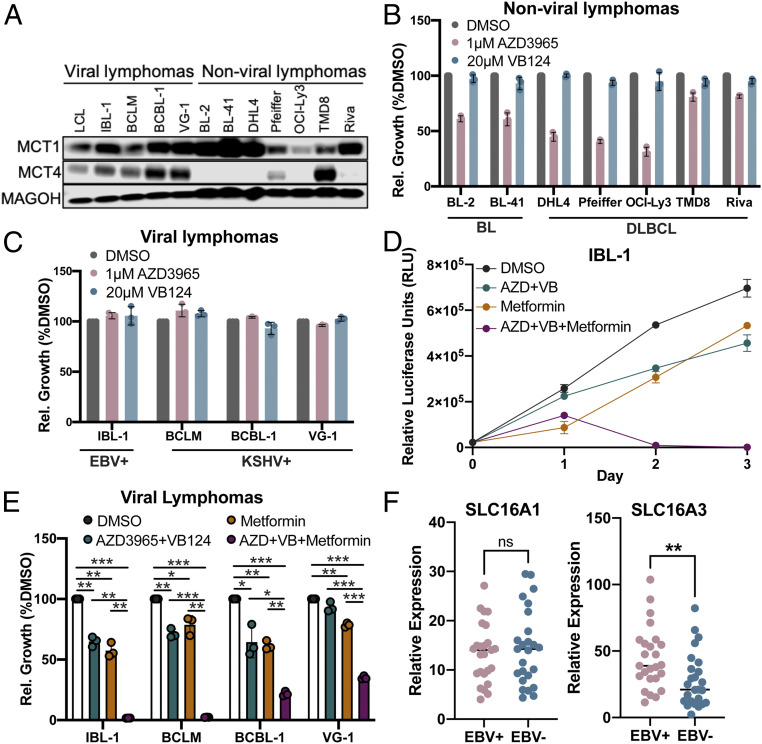
Epstein-Barr virus (EBV) is a ubiquitous herpesvirus that typically causes asymptomatic infection but can promote B lymphoid tumors in the immune suppressed. In vitro, EBV infection of primary B cells stimulates glycolysis during immortalization into lymphoblastoid cell lines (LCLs). Lactate export during glycolysis is crucial for continued proliferation of many cancer cells-part of a phenomenon known as the "Warburg effect"- and is mediated by monocarboxylate transporters (MCTs). However, the role of MCTs has yet to be studied in EBV-associated malignancies, which display Warburg-like metabolism in vitro. Here, we show that EBV infection of B lymphocytes directly promotes temporal induction of MCT1 and MCT4 through the viral proteins EBNA2 and LMP1, respectively. Functionally, MCT1 was required for early B cell proliferation, and MCT4 up-regulation promoted acquired resistance to MCT1 antagonism in LCLs. However, dual MCT1/4 inhibition led to LCL growth arrest and lactate buildup. Metabolic profiling in LCLs revealed significantly reduced oxygen consumption rates (OCRs) and NAD+/NADH ratios, contrary to previous observations of increased OCR and unaltered NAD+/NADH ratios in MCT1/4-inhibited cancer cells. Furthermore, U-13C6-glucose labeling of MCT1/4-inhibited LCLs revealed depleted glutathione pools that correlated with elevated reactive oxygen species. Finally, we found that dual MCT1/4 inhibition also sensitized LCLs to killing by the electron transport chain complex I inhibitors phenformin and metformin. These findings were extended to viral lymphomas associated with EBV and the related gammaherpesvirus KSHV, pointing at a therapeutic approach for targeting both viral lymphomas.
Keywords: Epstein–Barr virus; cancer metabolism; lactate export; monocarboxylate transporter; viral lymphoma.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Babcock G. J., Decker L. L., Volk M., Thorley-Lawson D. A., EBV persistence in memory B cells in vivo. Immunity 9, 395–404 (1998). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
