

Fever supports CD8+ effector T cell responses by promoting mitochondrial translation
- PMID: 34161266
- PMCID: PMC8237659
- DOI: 10.1073/pnas.2023752118
Fever supports CD8+ effector T cell responses by promoting mitochondrial translation
Abstract
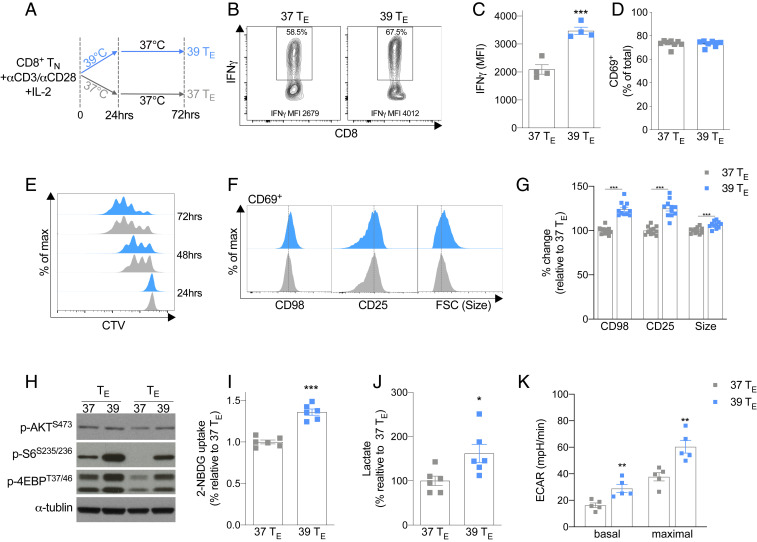
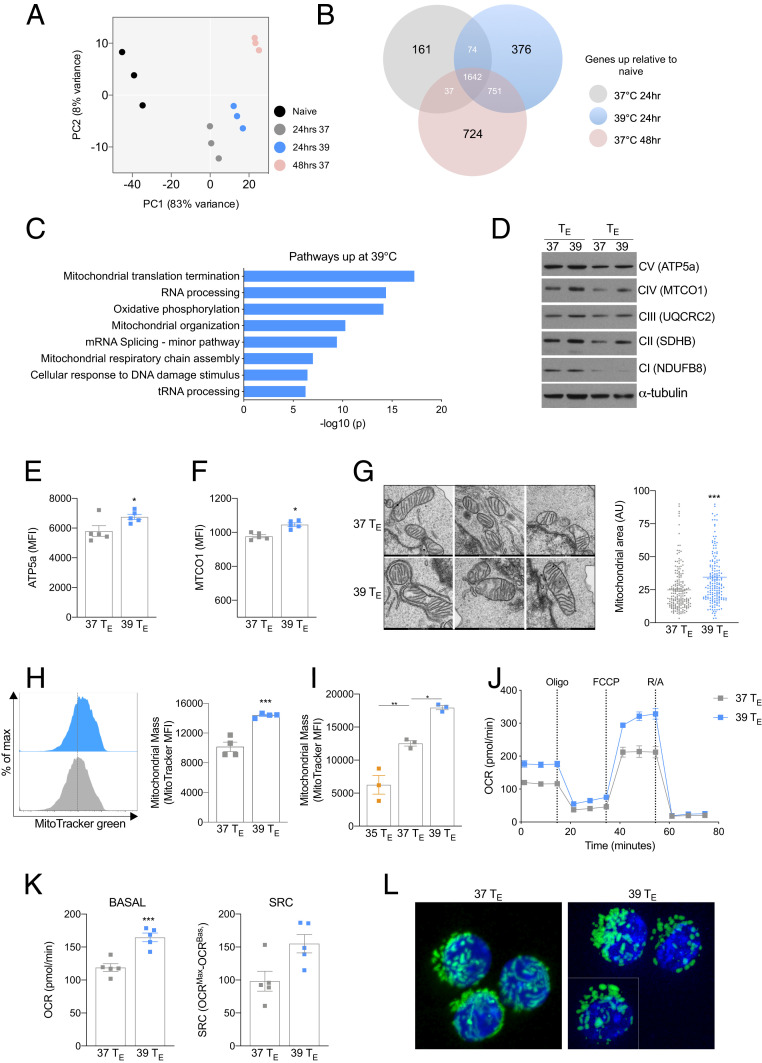
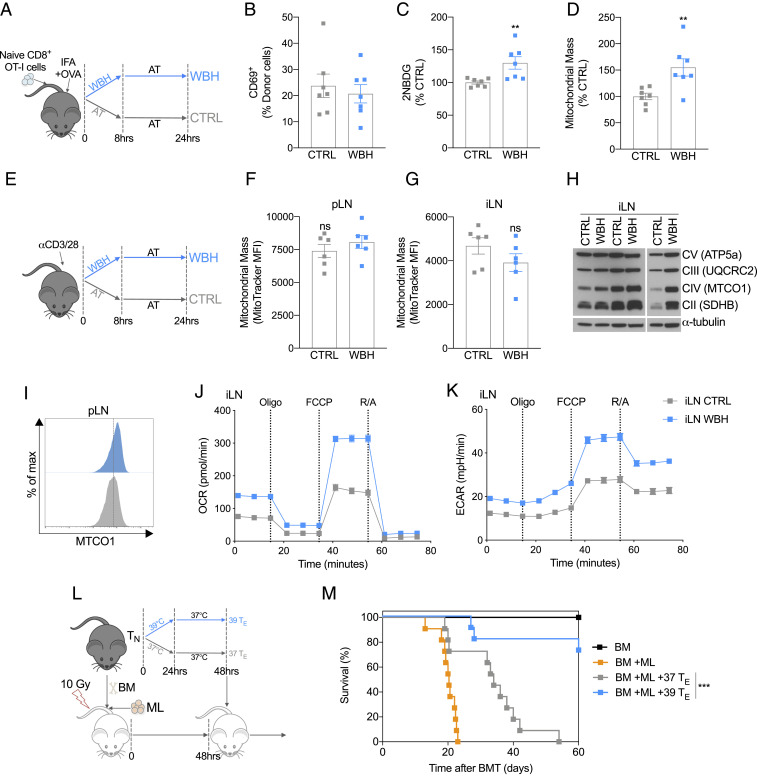
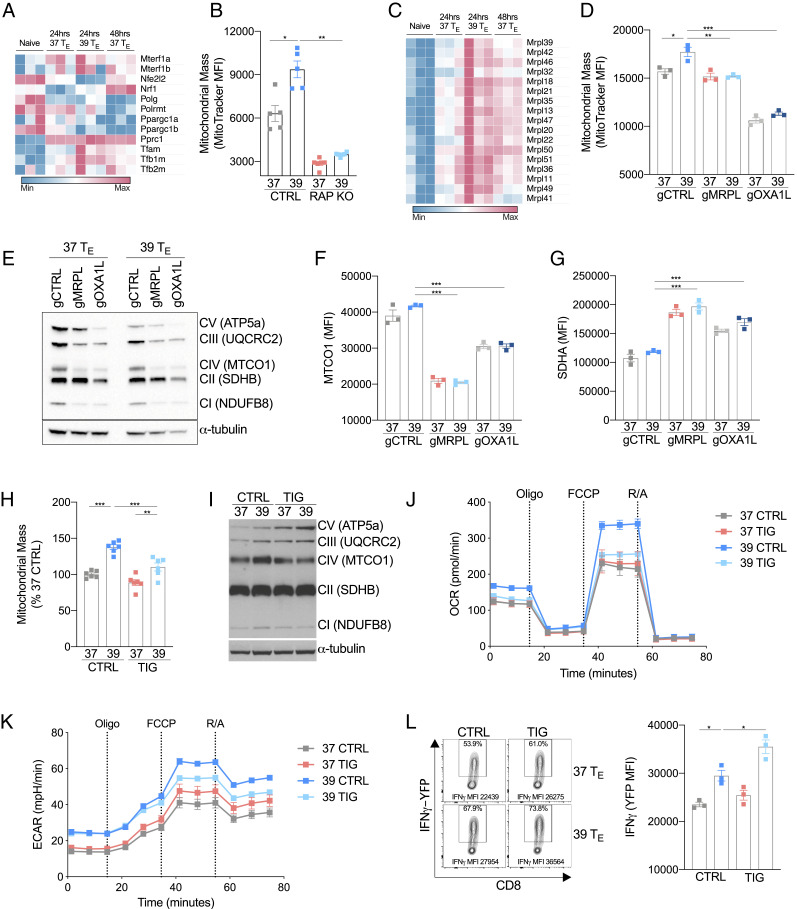
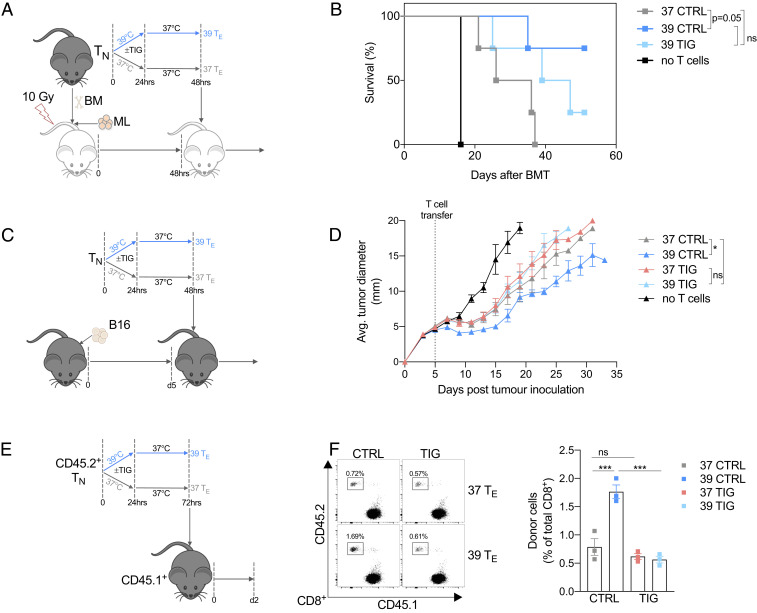
Fever can provide a survival advantage during infection. Metabolic processes are sensitive to environmental conditions, but the effect of fever on T cell metabolism is not well characterized. We show that in activated CD8+ T cells, exposure to febrile temperature (39 °C) augmented metabolic activity and T cell effector functions, despite having a limited effect on proliferation or activation marker expression. Transcriptional profiling revealed an up-regulation of mitochondrial pathways, which was consistent with increased mass and metabolism observed in T cells exposed to 39 °C. Through in vitro and in vivo models, we determined that mitochondrial translation is integral to the enhanced metabolic activity and function of CD8+ T cells exposed to febrile temperature. Transiently exposing donor lymphocytes to 39 °C prior to infusion in a myeloid leukemia mouse model conferred enhanced therapeutic efficacy, raising the possibility that exposure of T cells to febrile temperatures could have clinical potential.
Keywords: T cell; fever; immunology; metabolism; mitochondria.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
Competing interest statement: E.L.P. is a Scientific Advisory Board member of ImmunoMet Therapeutics, and E.J.P. and E.L.P. are founders of Rheos Medicines.
Figures
References
-
- Wang X., et al. ., Febrile temperature critically controls the differentiation and pathogenicity of T helper 17 cells. Immunity 52, 328–341.e5 (2020). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
