

Lactate and IL6 define separable paths of inflammatory metabolic adaptation
- PMID: 34162546
- PMCID: PMC8221612
- DOI: 10.1126/sciadv.abg3505
Lactate and IL6 define separable paths of inflammatory metabolic adaptation
Abstract
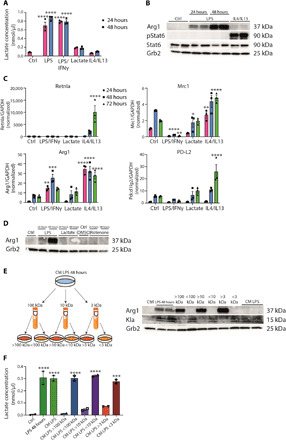
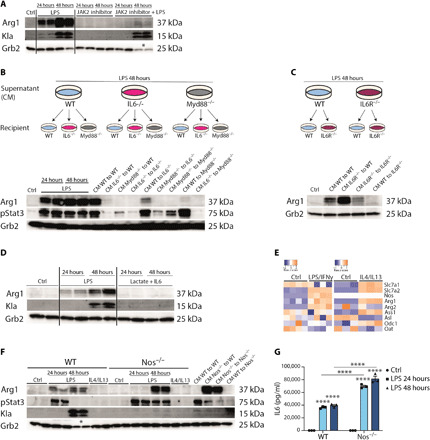
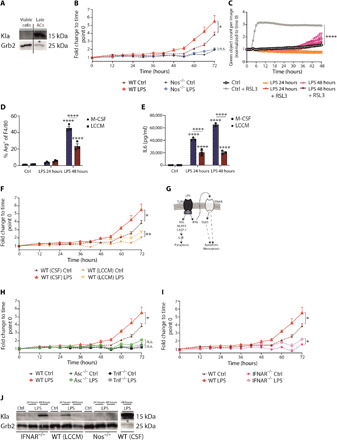
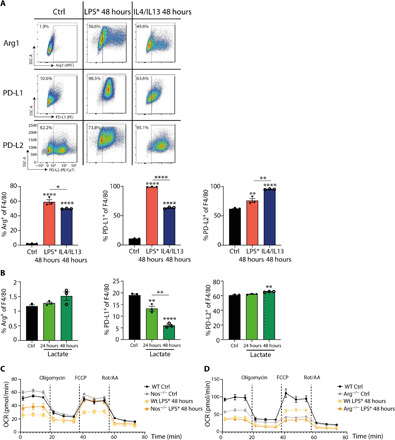
Lactate is an end point of Warburg-type metabolism found in inflammatory macrophages. Recently, lactate was shown to modify histones of lipopolysaccharide (LPS)-activated macrophages in a time-dependent way and promote the expression of genes linked to tissue repair, including arginase-1 (Arg1). We tested the interrelationships between histone lactylation (Kla) and tissue reparative gene expression and found that Kla was uncoupled from changes in gene expression linked to resolving M2 macrophage activation but correlated with Arg1 expression. LPS-induced Arg1 was instead dependent on autocrine-paracrine interleukin-6 (IL6) production, the IL6 receptor, and Stat3 signal transduction. We found that Kla increases as macrophages prepare to die under inflammatory stress, and Kla was absent in macrophages that cannot generate reactive nitrogen or have defects in diverse macrophage death pathways. Thus, Kla is a consequence rather than a cause of macrophage activation but occurs coincidently with an IL6- and Arg1-dependent metabolic rewiring under inflammatory duress.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Zhang D., Tang Z., Huang H., Zhou G., Cui C., Weng Y., Liu W., Kim S., Lee S., Perez-Neut M., Ding J., Czyz D., Hu R., Ye Z., He M., Zheng Y. G., Shuman H. A., Dai L., Ren B., Roeder R. G., Becker L., Zhao Y., Metabolic regulation of gene expression by histone lactylation. Nature 574, 575–580 (2019). - PMC - PubMed
-
- Diskin C., Ryan T. A. J., O’Neill L. A. J., Modification of proteins by metabolites in immunity. Immunity 54, 19–31 (2021). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
