

Immunodominant T-cell epitopes from the SARS-CoV-2 spike antigen reveal robust pre-existing T-cell immunity in unexposed individuals
- PMID: 34162945
- PMCID: PMC8222233
- DOI: 10.1038/s41598-021-92521-4
Immunodominant T-cell epitopes from the SARS-CoV-2 spike antigen reveal robust pre-existing T-cell immunity in unexposed individuals
Abstract
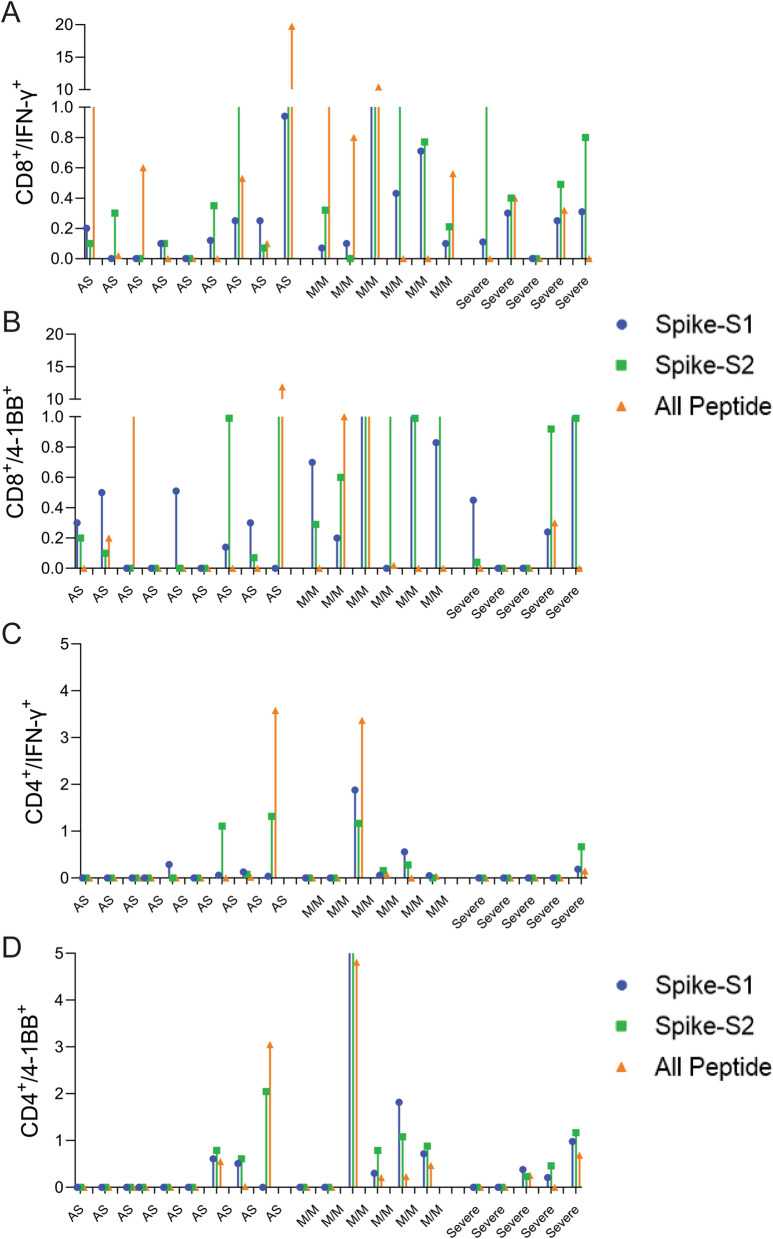
The COVID-19 pandemic has revealed a range of disease phenotypes in infected patients with asymptomatic, mild, or severe clinical outcomes, but the mechanisms that determine such variable outcomes remain unresolved. In this study, we identified immunodominant CD8 T-cell epitopes in the spike antigen using a novel TCR-binding algorithm. The predicted epitopes induced robust T-cell activation in unexposed donors demonstrating pre-existing CD4 and CD8 T-cell immunity to SARS-CoV-2 antigen. The T-cell reactivity to the predicted epitopes was higher than the Spike-S1 and S2 peptide pools in the unexposed donors. A key finding of our study is that pre-existing T-cell immunity to SARS-CoV-2 is contributed by TCRs that recognize common viral antigens such as Influenza and CMV, even though the viral epitopes lack sequence identity to the SARS-CoV-2 epitopes. This finding is in contrast to multiple published studies in which pre-existing T-cell immunity is suggested to arise from shared epitopes between SARS-CoV-2 and other common cold-causing coronaviruses. However, our findings suggest that SARS-CoV-2 reactive T-cells are likely to be present in many individuals because of prior exposure to flu and CMV viruses.
Conflict of interest statement
All authors are full-time employees of MedGenome Inc. 348 Hatch Drive, Foster City, CA 94404, USA or MedGenome Labs Limited, 258/A 3rd Floor, Narayana Nethralaya, Hosur Road, Bangalore 560099, India and receive salary and benefits.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
