

Mechanism of disease and therapeutic rescue of Dok7 congenital myasthenia
- PMID: 34163073
- PMCID: PMC8277574
- DOI: 10.1038/s41586-021-03672-3
Mechanism of disease and therapeutic rescue of Dok7 congenital myasthenia
Abstract
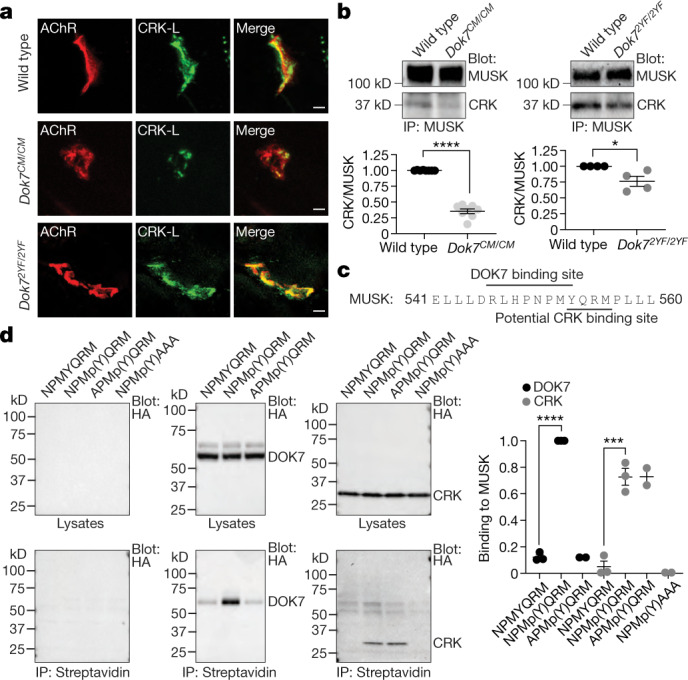
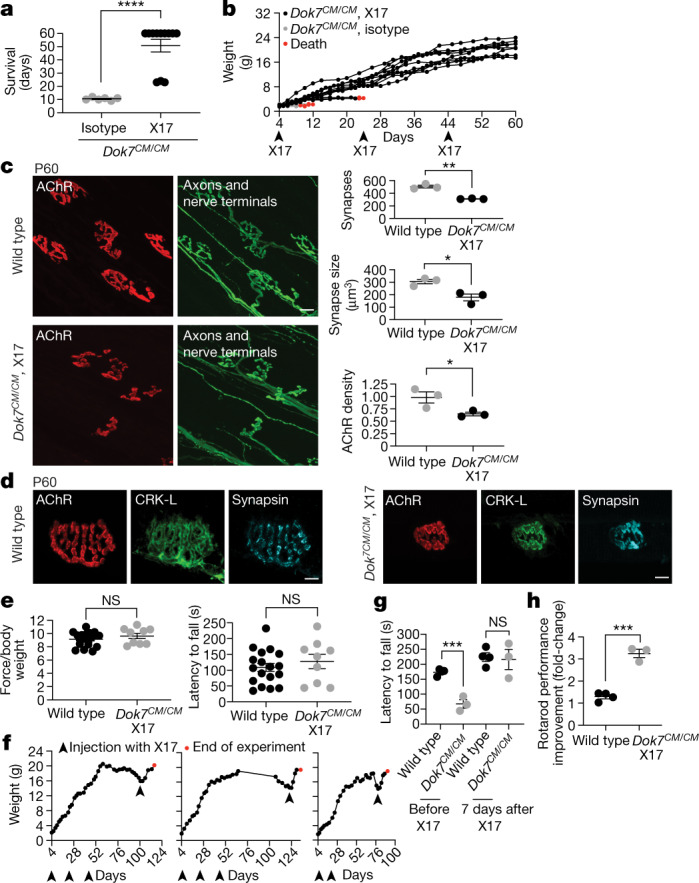
Congenital myasthenia (CM) is a devastating neuromuscular disease, and mutations in DOK7, an adaptor protein that is crucial for forming and maintaining neuromuscular synapses, are a major cause of CM1,2. The most common disease-causing mutation (DOK71124_1127 dup) truncates DOK7 and leads to the loss of two tyrosine residues that are phosphorylated and recruit CRK proteins, which are important for anchoring acetylcholine receptors at synapses. Here we describe a mouse model of this common form of CM (Dok7CM mice) and a mouse with point mutations in the two tyrosine residues (Dok72YF). We show that Dok7CM mice had severe deficits in neuromuscular synapse formation that caused neonatal lethality. Unexpectedly, these deficits were due to a severe deficiency in phosphorylation and activation of muscle-specific kinase (MUSK) rather than a deficiency in DOK7 tyrosine phosphorylation. We developed agonist antibodies against MUSK and show that these antibodies restored neuromuscular synapse formation and prevented neonatal lethality and late-onset disease in Dok7CM mice. These findings identify an unexpected cause for disease and a potential therapy for both DOK7 CM and other forms of CM caused by mutations in AGRIN, LRP4 or MUSK, and illustrate the potential of targeted therapy to rescue congenital lethality.
© 2021. The Author(s).
Conflict of interest statement
S.J.B. is an inventor on a patent (no. 9,329,182) for ‘Method of treating motor neuron disease with an antibody that agonizes MUSK’. S.J.B., S.K., J.O., A.K. and N.L. are inventors on a patent application (no. 1474662.02232) for ‘Therapeutic MUSK Antibodies’ filed by New York University. These patents have been licensed to Argenx BVBA. S.J.B. and S.K. received research funding from Argenx BVBA. S.K. is a scientific advisory board member and holds equity in and receives consulting fees from Black Diamond Therapeutics, and receives research funding from Puretech Health.
Figures










Comment in
-
Docking downstream to treat myasthenia.Nat Rev Drug Discov. 2021 Aug;20(8):587. doi: 10.1038/d41573-021-00120-9. Nat Rev Drug Discov. 2021. PMID: 34239091 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
