

Identifying the combinatorial control of signal-dependent transcription factors
- PMID: 34166361
- PMCID: PMC8263068
- DOI: 10.1371/journal.pcbi.1009095
Identifying the combinatorial control of signal-dependent transcription factors
Abstract
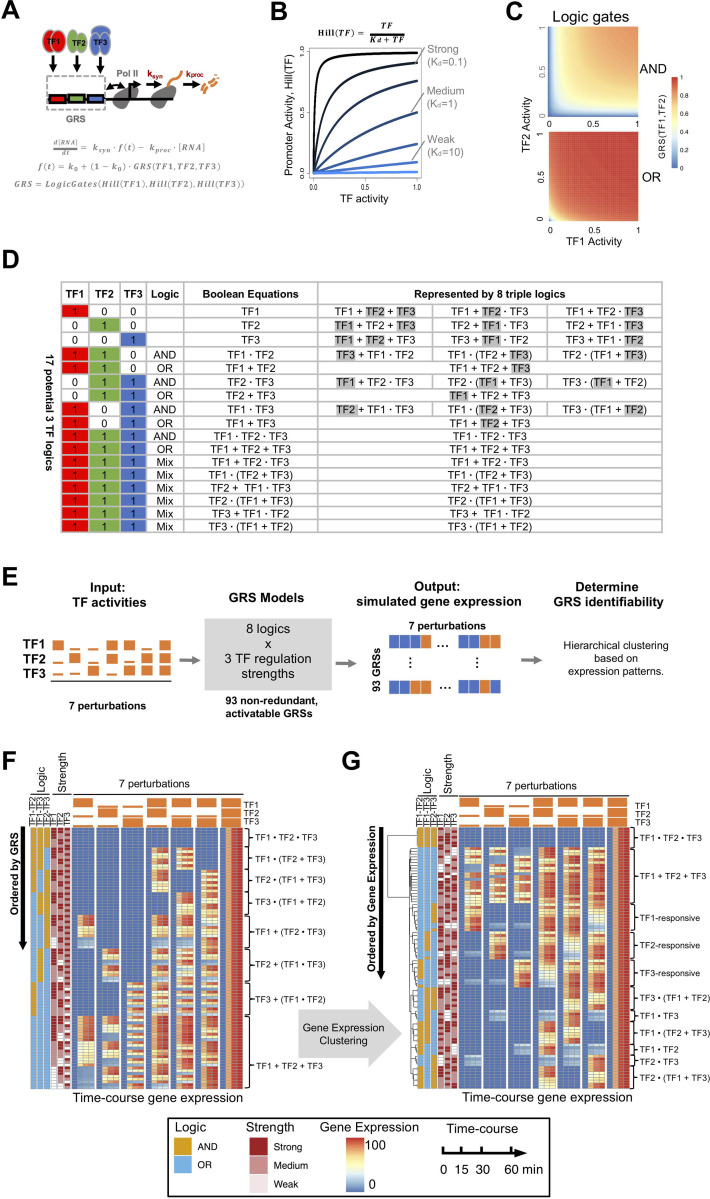
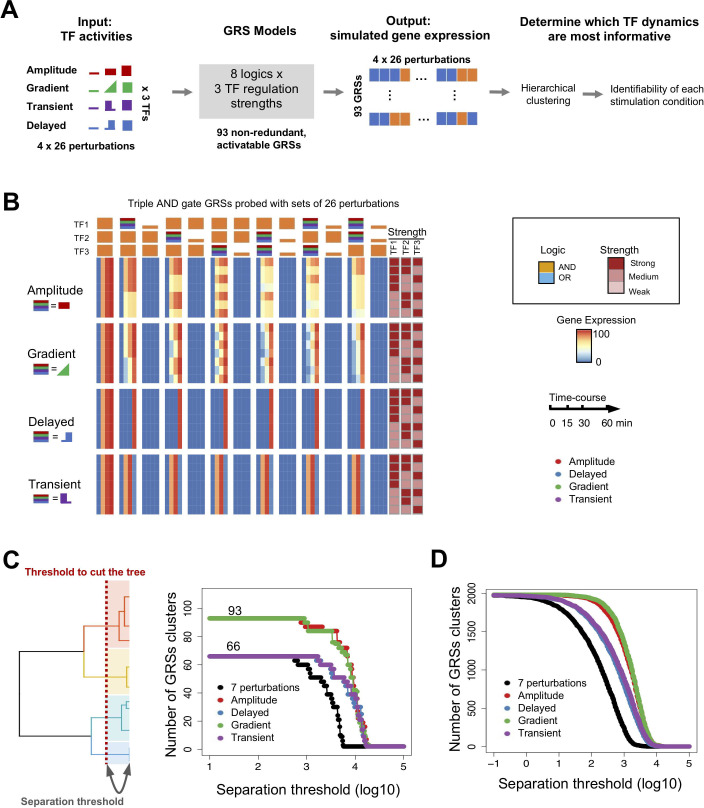
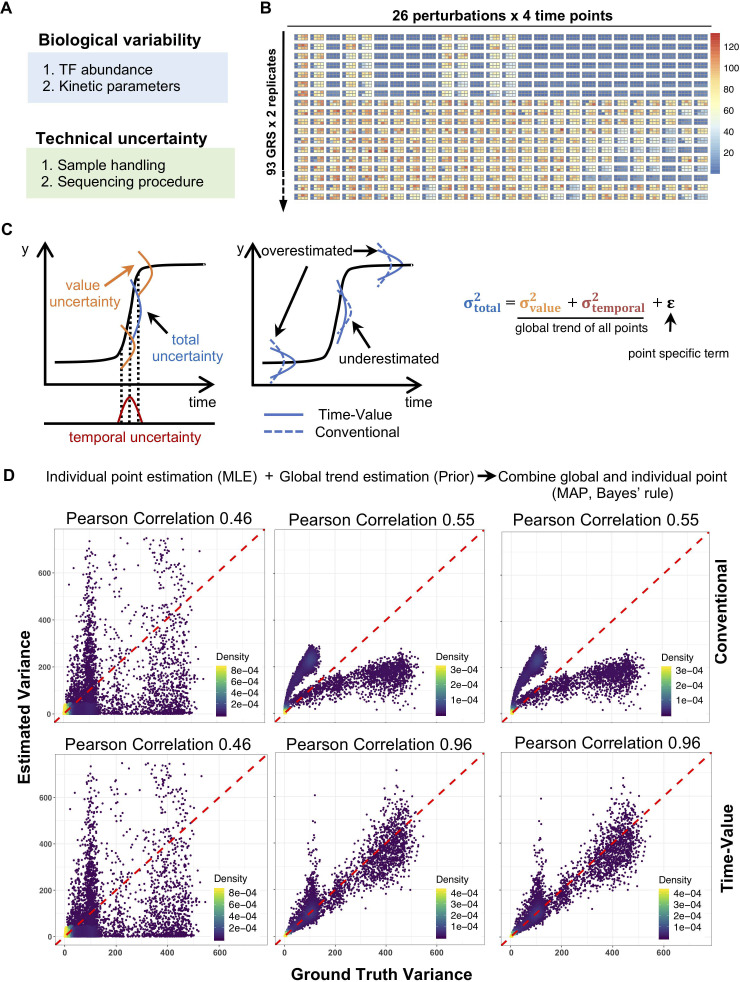
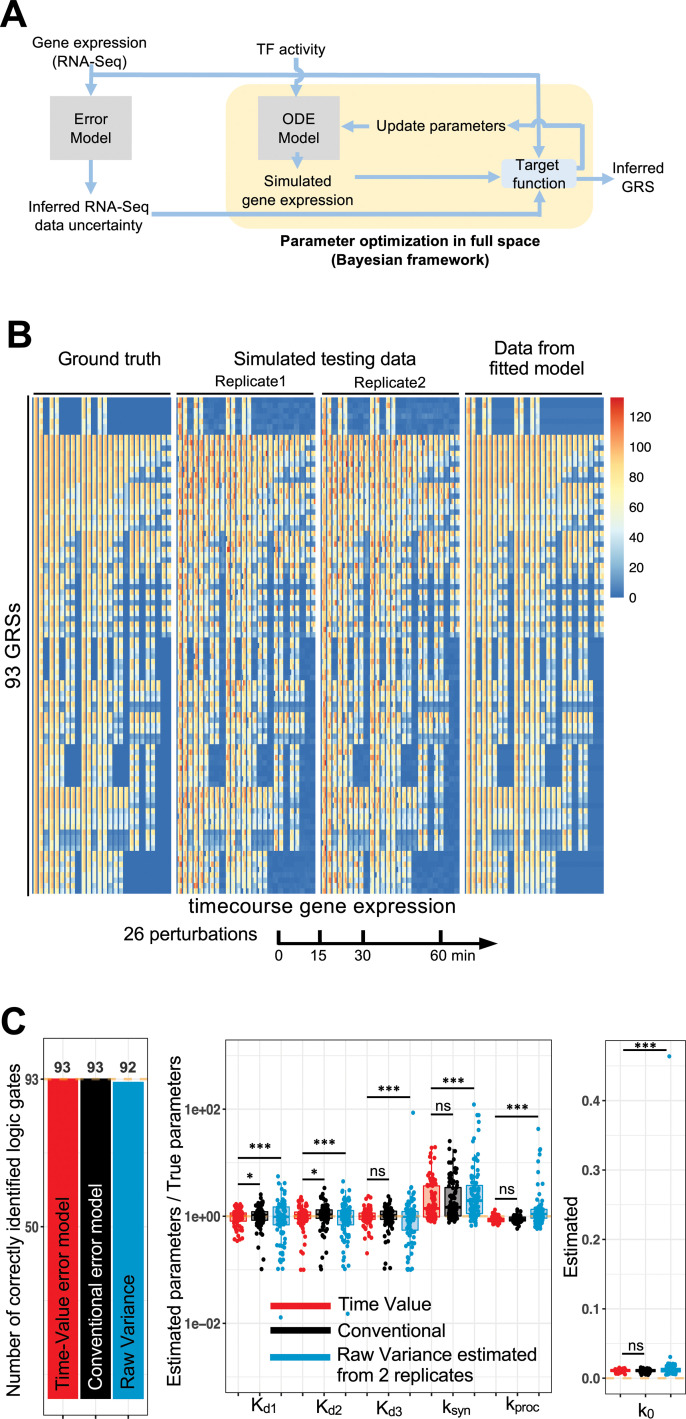
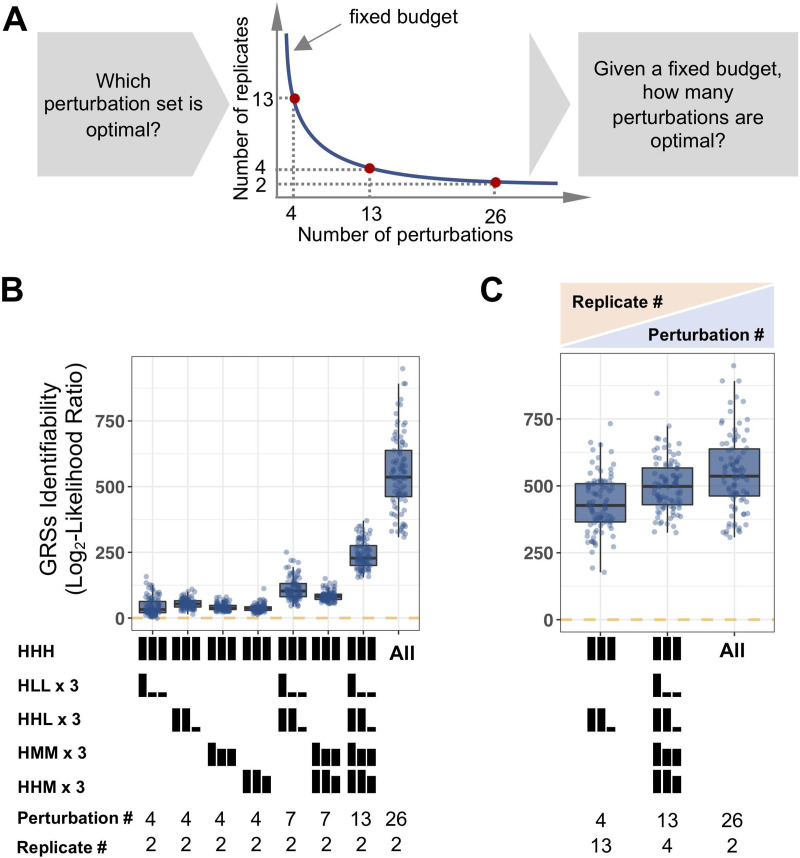
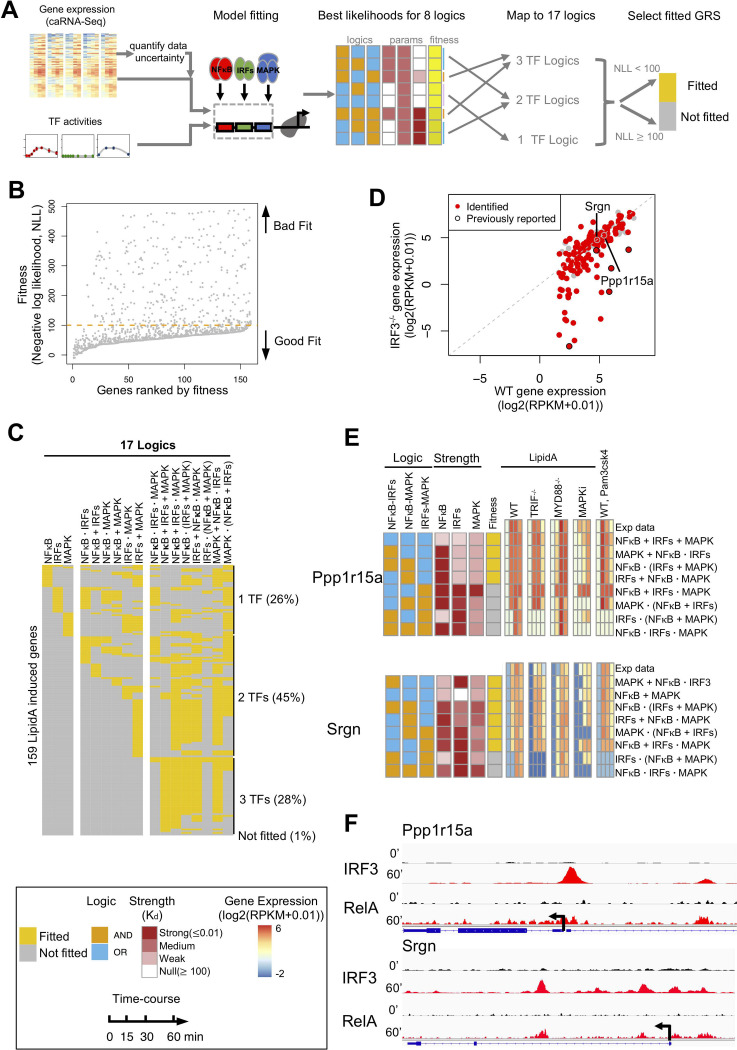
The effectiveness of immune responses depends on the precision of stimulus-responsive gene expression programs. Cells specify which genes to express by activating stimulus-specific combinations of stimulus-induced transcription factors (TFs). Their activities are decoded by a gene regulatory strategy (GRS) associated with each response gene. Here, we examined whether the GRSs of target genes may be inferred from stimulus-response (input-output) datasets, which remains an unresolved model-identifiability challenge. We developed a mechanistic modeling framework and computational workflow to determine the identifiability of all possible combinations of synergistic (AND) or non-synergistic (OR) GRSs involving three transcription factors. Considering different sets of perturbations for stimulus-response studies, we found that two thirds of GRSs are easily distinguishable but that substantially more quantitative data is required to distinguish the remaining third. To enhance the accuracy of the inference with timecourse experimental data, we developed an advanced error model that avoids error overestimates by distinguishing between value and temporal error. Incorporating this error model into a Bayesian framework, we show that GRS models can be identified for individual genes by considering multiple datasets. Our analysis rationalizes the allocation of experimental resources by identifying most informative TF stimulation conditions. Applying this computational workflow to experimental data of immune response genes in macrophages, we found that a much greater fraction of genes are combinatorially controlled than previously reported by considering compensation among transcription factors. Specifically, we revealed that a group of known NFκB target genes may also be regulated by IRF3, which is supported by chromatin immuno-precipitation analysis. Our study provides a computational workflow for designing and interpreting stimulus-response gene expression studies to identify underlying gene regulatory strategies and further a mechanistic understanding.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Salleh FH, Arif SM, Zainudin S, Firdaus-Raih M. Reconstructing gene regulatory networks from knock-out data using Gaussian Noise Model and Pearson Correlation Coefficient. Computational biology and chemistry. 2015. Dec 1;59:3–14. Epub 2015 Jun 17. doi: 10.1016/j.compbiolchem.2015.04.012 . - DOI - PubMed
-
- Margolin AA, Nemenman I, Basso K, Wiggins C, Stolovitzky G, Dalla Favera R, et al.. ARACNE: an algorithm for the reconstruction of gene regulatory networks in a mammalian cellular context. BMC bioinformatics 2006. Mar (Vol. 7, No. 1, pp. 1–15). BioMed Central. doi: 10.1186/1471-2105-7-S1-S7 ; PMCID: PMC1810318. - DOI - PMC - PubMed
-
- Faith JJ, Hayete B, Thaden JT, Mogno I, Wierzbowski J, Cottarel G, et al.. Large-scale mapping and validation of Escherichia coli transcriptional regulation from a compendium of expression profiles. PLoS biol. 2007. Jan 9;5(1):e8. doi: 10.1371/journal.pbio.0050008 ; PMCID: PMC1764438. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
