

SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity
- PMID: 34171266
- PMCID: PMC8205251
- DOI: 10.1016/j.chom.2021.06.006
SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity
Abstract
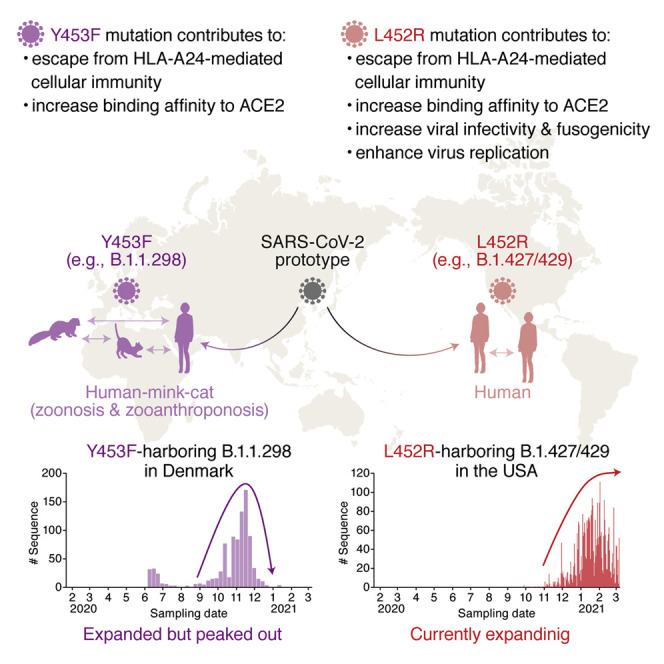
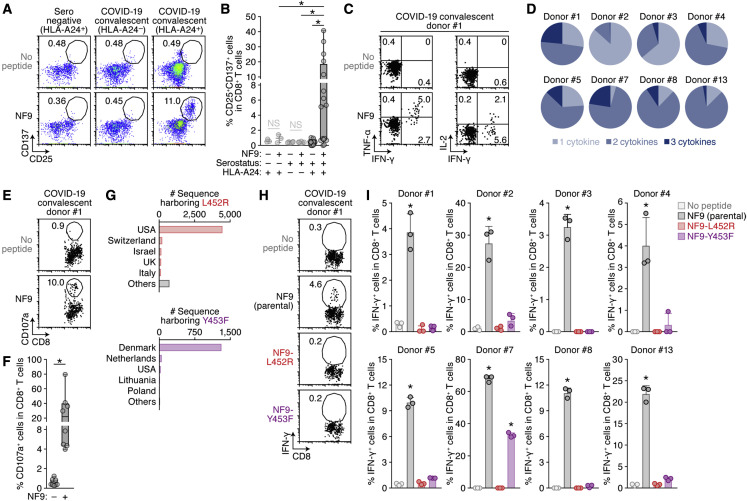
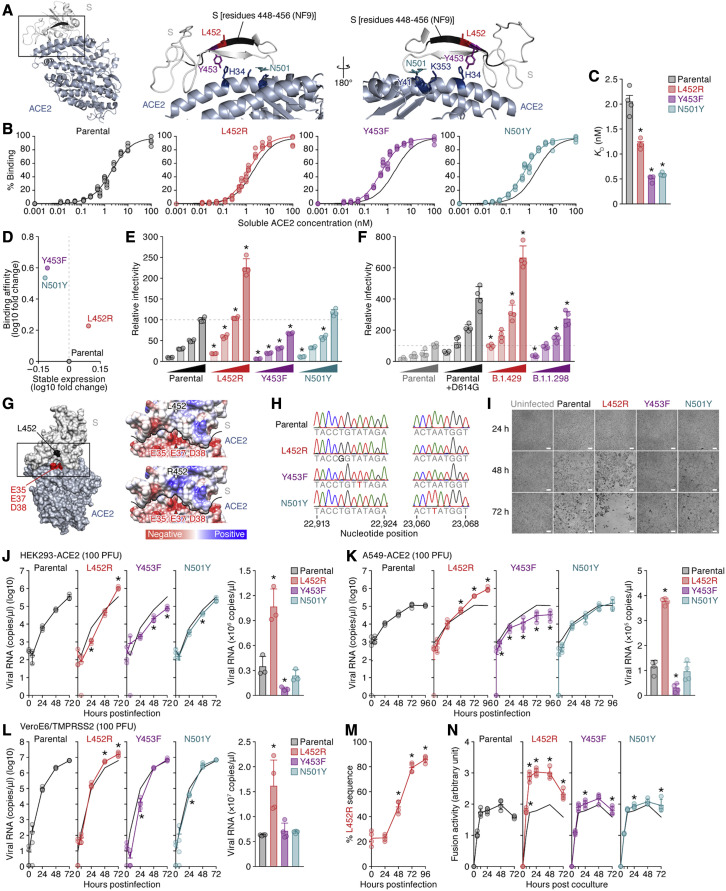
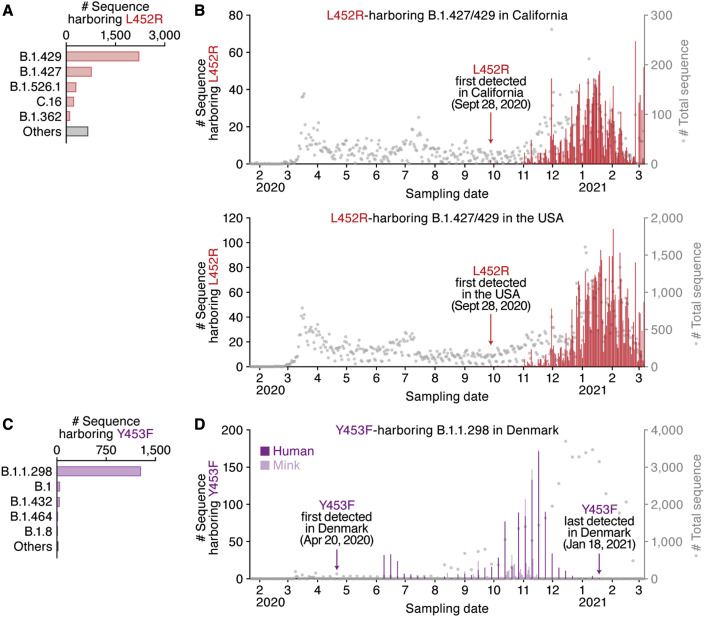
Many SARS-CoV-2 variants with naturally acquired mutations have emerged. These mutations can affect viral properties such as infectivity and immune resistance. Although the sensitivity of naturally occurring SARS-CoV-2 variants to humoral immunity has been investigated, sensitivity to human leukocyte antigen (HLA)-restricted cellular immunity remains largely unexplored. Here, we demonstrate that two recently emerging mutations in the receptor-binding domain of the SARS-CoV-2 spike protein, L452R (in B.1.427/429 and B.1.617) and Y453F (in B.1.1.298), confer escape from HLA-A24-restricted cellular immunity. These mutations reinforce affinity toward the host entry receptor ACE2. Notably, the L452R mutation increases spike stability, viral infectivity, viral fusogenicity, and thereby promotes viral replication. These data suggest that HLA-restricted cellular immunity potentially affects the evolution of viral phenotypes and that a further threat of the SARS-CoV-2 pandemic is escape from cellular immunity.
Keywords: B.1.1.298; B.1.427/429; COVID-19; L452R; SARS-CoV-2; Y453F; cellular immunity; naturally occurring variants; receptor-binding motif; spike protein.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
