

Structural basis of transcriptional activation by the Mycobacterium tuberculosis intrinsic antibiotic-resistance transcription factor WhiB7
- PMID: 34171296
- PMCID: PMC8311663
- DOI: 10.1016/j.molcel.2021.05.017
Structural basis of transcriptional activation by the Mycobacterium tuberculosis intrinsic antibiotic-resistance transcription factor WhiB7
Abstract
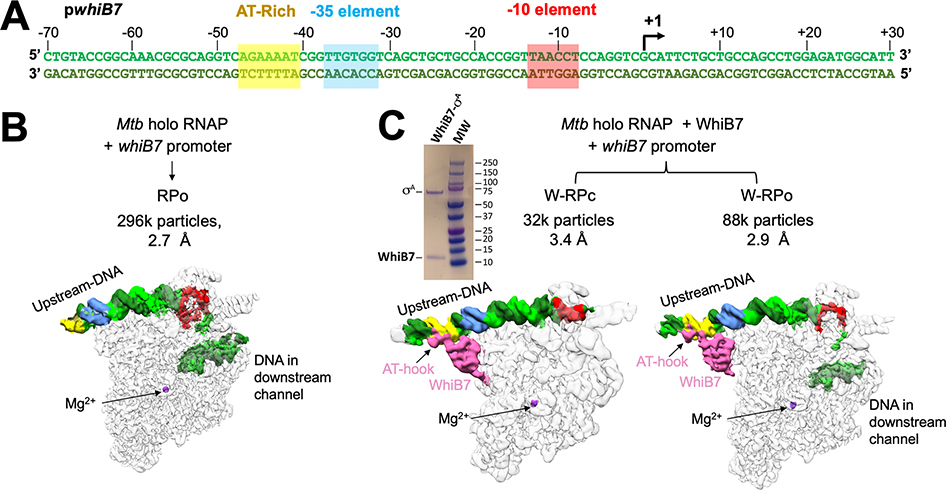
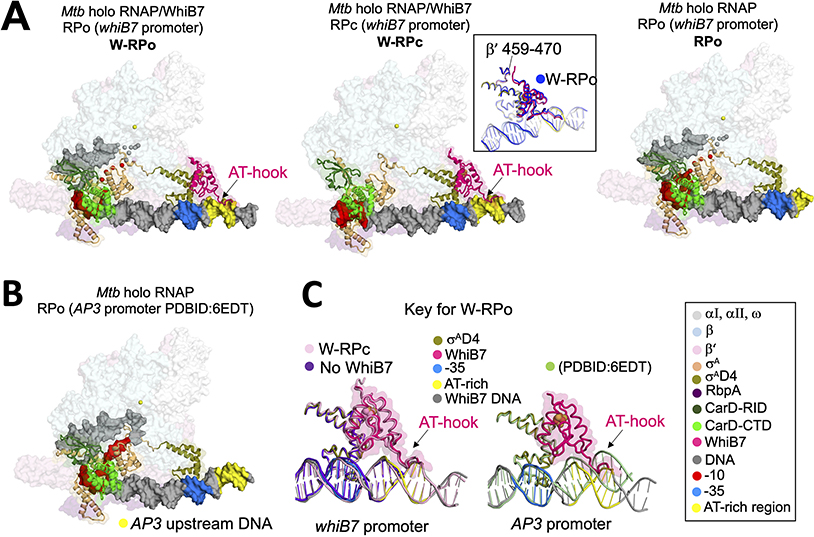
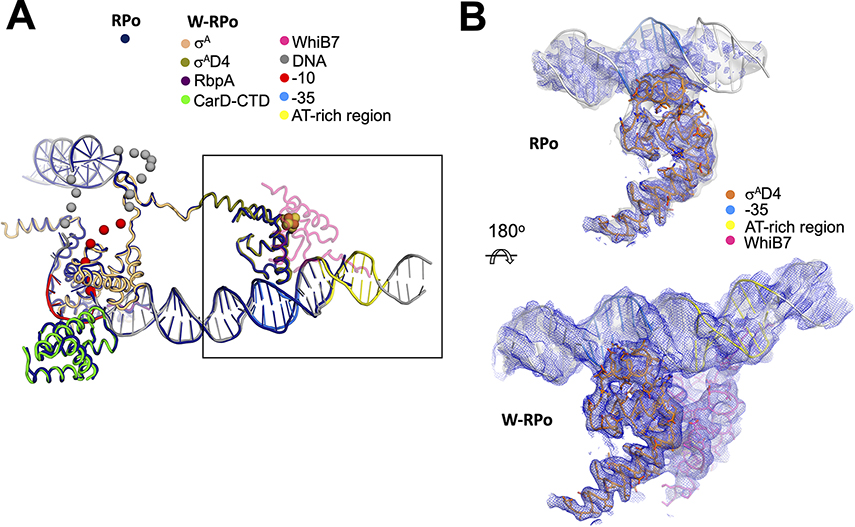
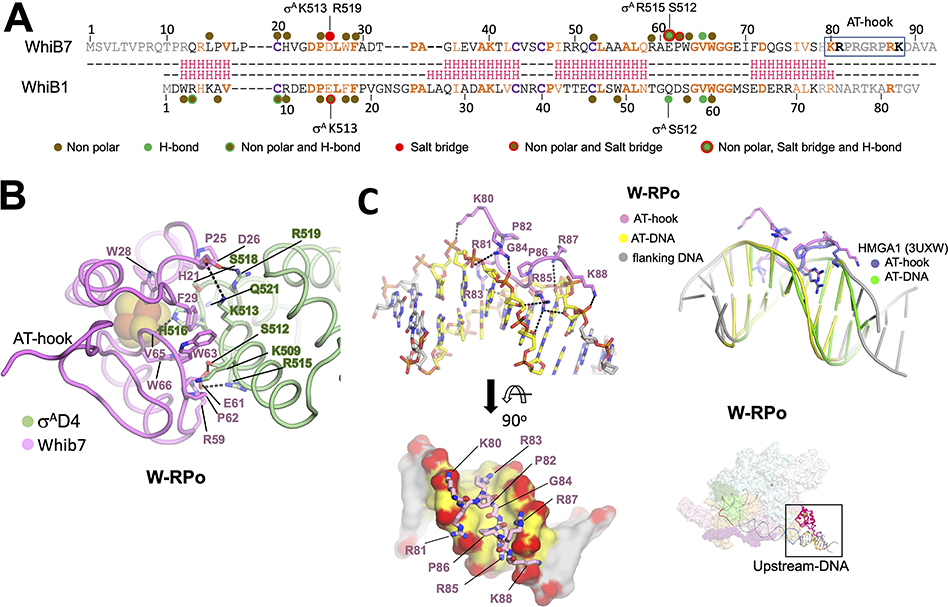
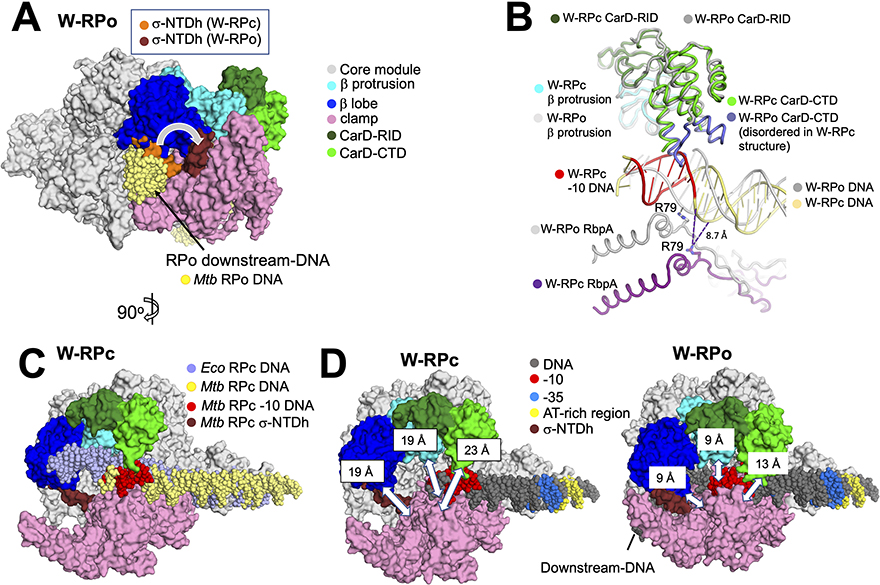
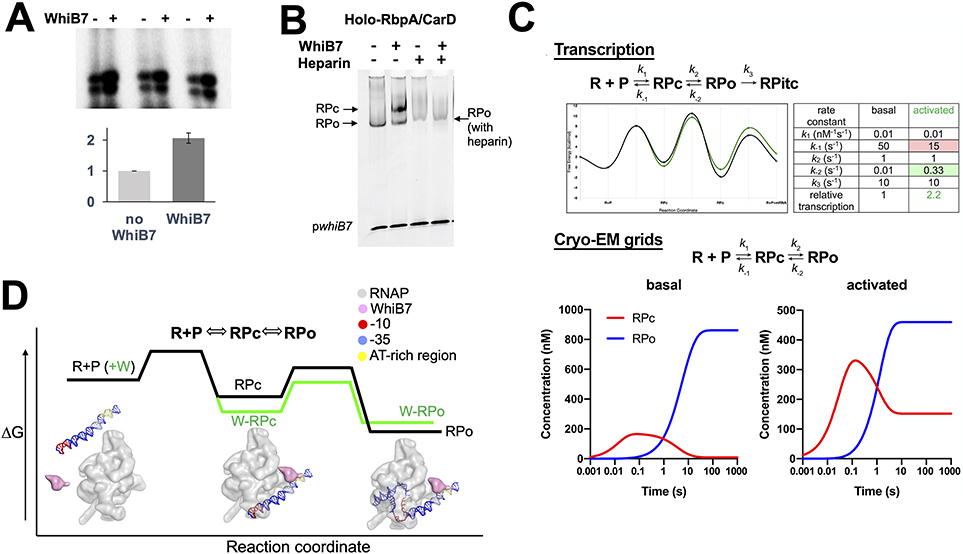
In pathogenic mycobacteria, transcriptional responses to antibiotics result in induced antibiotic resistance. WhiB7 belongs to the Actinobacteria-specific family of Fe-S-containing transcription factors and plays a crucial role in inducible antibiotic resistance in mycobacteria. Here, we present cryoelectron microscopy structures of Mycobacterium tuberculosis transcriptional regulatory complexes comprising RNA polymerase σA-holoenzyme, global regulators CarD and RbpA, and WhiB7, bound to a WhiB7-regulated promoter. The structures reveal how WhiB7 interacts with σA-holoenzyme while simultaneously interacting with an AT-rich sequence element via its AT-hook. Evidently, AT-hooks, rare elements in bacteria yet prevalent in eukaryotes, bind to target AT-rich DNA sequences similarly to the nuclear chromosome binding proteins. Unexpectedly, a subset of particles contained a WhiB7-stabilized closed promoter complex, revealing this intermediate's structure, and we apply kinetic modeling and biochemical assays to rationalize how WhiB7 activates transcription. Altogether, our work presents a comprehensive view of how WhiB7 serves to activate gene expression leading to antibiotic resistance.
Keywords: RNA polymerase; WhiB7; antibiotic resistance; cryo-EM; iron cluster; transcription; transcription factor; transcription initiation.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Structural insights into the functional divergence of WhiB-like proteins in Mycobacterium tuberculosis.Mol Cell. 2021 Jul 15;81(14):2887-2900.e5. doi: 10.1016/j.molcel.2021.06.002. Epub 2021 Jun 24. Mol Cell. 2021. PMID: 34171298 Free PMC article.
-
WhiB7, an Fe-S-dependent transcription factor that activates species-specific repertoires of drug resistance determinants in actinobacteria.J Biol Chem. 2013 Nov 29;288(48):34514-28. doi: 10.1074/jbc.M113.516385. Epub 2013 Oct 14. J Biol Chem. 2013. PMID: 24126912 Free PMC article.
-
The WblC/WhiB7 Transcription Factor Controls Intrinsic Resistance to Translation-Targeting Antibiotics by Altering Ribosome Composition.mBio. 2020 Apr 14;11(2):e00625-20. doi: 10.1128/mBio.00625-20. mBio. 2020. PMID: 32291305 Free PMC article.
-
WhiB7, a transcriptional activator that coordinates physiology with intrinsic drug resistance in Mycobacterium tuberculosis.Expert Rev Anti Infect Ther. 2012 Sep;10(9):1037-47. doi: 10.1586/eri.12.90. Expert Rev Anti Infect Ther. 2012. PMID: 23106278 Review.
-
SigR, a hub of multilayered regulation of redox and antibiotic stress responses.Mol Microbiol. 2019 Aug;112(2):420-431. doi: 10.1111/mmi.14341. Epub 2019 Jul 26. Mol Microbiol. 2019. PMID: 31269533 Review.
Cited by
-
Early intermediates in bacterial RNA polymerase promoter melting visualized by time-resolved cryo-electron microscopy.Nat Struct Mol Biol. 2024 Nov;31(11):1778-1788. doi: 10.1038/s41594-024-01349-9. Epub 2024 Jul 1. Nat Struct Mol Biol. 2024. PMID: 38951624
-
A Feedback Regulatory Loop Containing McdR and WhiB2 Controls Cell Division and DNA Repair in Mycobacteria.mBio. 2022 Apr 26;13(2):e0334321. doi: 10.1128/mbio.03343-21. Epub 2022 Mar 31. mBio. 2022. PMID: 35357209 Free PMC article.
-
Structural basis of three different transcription activation strategies adopted by a single regulator SoxS.Nucleic Acids Res. 2022 Oct 28;50(19):11359-11373. doi: 10.1093/nar/gkac898. Nucleic Acids Res. 2022. PMID: 36243985 Free PMC article.
-
WhiB-like proteins: Diversity of structure, function and mechanism.Biochim Biophys Acta Mol Cell Res. 2024 Oct;1871(7):119787. doi: 10.1016/j.bbamcr.2024.119787. Epub 2024 Jun 13. Biochim Biophys Acta Mol Cell Res. 2024. PMID: 38879133 Review.
-
Early intermediates in bacterial RNA polymerase promoter melting visualized by time-resolved cryo-electron microscopy.bioRxiv [Preprint]. 2024 Mar 14:2024.03.13.584744. doi: 10.1101/2024.03.13.584744. bioRxiv. 2024. Update in: Nat Struct Mol Biol. 2024 Nov;31(11):1778-1788. doi: 10.1038/s41594-024-01349-9. PMID: 38559232 Free PMC article. Updated. Preprint.
References
-
- Adams PD, Afonine PV, Bunkóczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung LW, Kapral GJ, Grosse-Kunstleve RW, et al. (2010). PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallographica Section D: Biological Crystallography 66, 213–221. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
