

Epigenetic clocks reveal a rejuvenation event during embryogenesis followed by aging
- PMID: 34172448
- PMCID: PMC8232908
- DOI: 10.1126/sciadv.abg6082
Epigenetic clocks reveal a rejuvenation event during embryogenesis followed by aging
Abstract
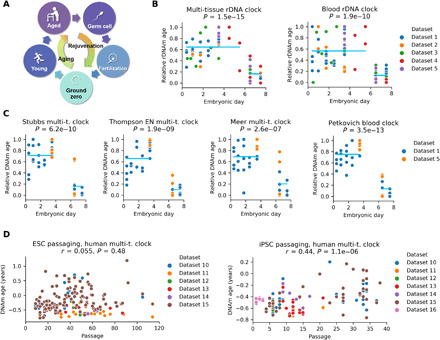
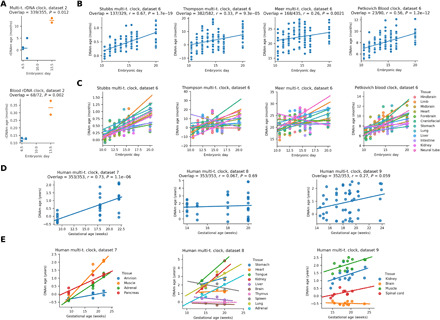
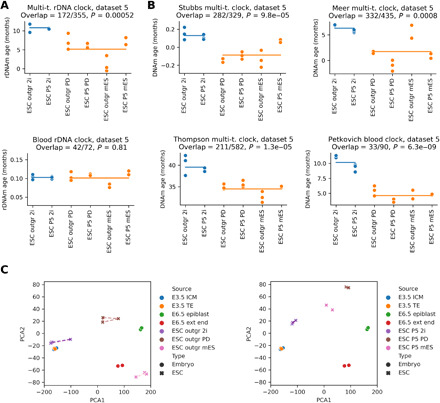
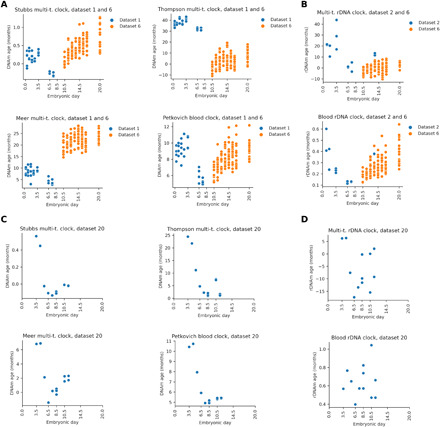
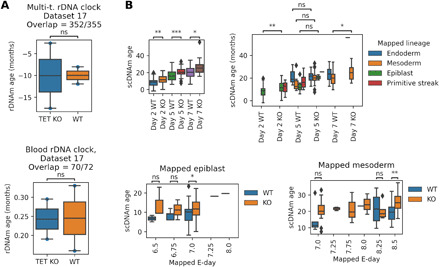
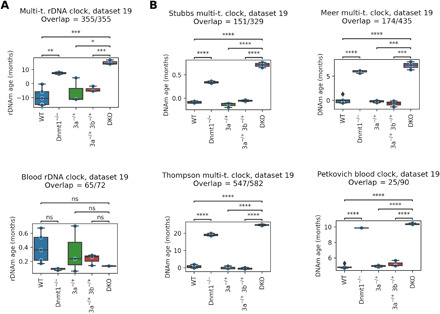
The notion that the germ line does not age goes back to the 19th-century ideas of August Weismann. However, being metabolically active, the germ line accumulates damage and other changes over time, i.e., it ages. For new life to begin in the same young state, the germ line must be rejuvenated in the offspring. Here, we developed a multi-tissue epigenetic clock and applied it, together with other aging clocks, to track changes in biological age during mouse and human prenatal development. This analysis revealed a significant decrease in biological age, i.e., rejuvenation, during early stages of embryogenesis, followed by an increase in later stages. We further found that pluripotent stem cells do not age even after extensive passaging and that the examined epigenetic age dynamics is conserved across species. Overall, this study uncovers a natural rejuvenation event during embryogenesis and suggests that the minimal biological age (ground zero) marks the beginning of organismal aging.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
Similar articles
-
Intersection clock reveals a rejuvenation event during human embryogenesis.Aging Cell. 2023 Oct;22(10):e13922. doi: 10.1111/acel.13922. Epub 2023 Oct 2. Aging Cell. 2023. PMID: 37786333 Free PMC article.
-
The Ground Zero of Organismal Life and Aging.Trends Mol Med. 2021 Jan;27(1):11-19. doi: 10.1016/j.molmed.2020.08.012. Epub 2020 Sep 23. Trends Mol Med. 2021. PMID: 32980264 Free PMC article. Review.
-
Rejuvenation by cell reprogramming: a new horizon in gerontology.Stem Cell Res Ther. 2018 Dec 17;9(1):349. doi: 10.1186/s13287-018-1075-y. Stem Cell Res Ther. 2018. PMID: 30558644 Free PMC article. Review.
-
Profiling epigenetic age in single cells.Nat Aging. 2021 Dec;1(12):1189-1201. doi: 10.1038/s43587-021-00134-3. Epub 2021 Dec 9. Nat Aging. 2021. PMID: 36211119 Free PMC article.
-
The Emerging View of Aging as a Reversible Epigenetic Process.Gerontology. 2017;63(5):426-431. doi: 10.1159/000477209. Epub 2017 May 25. Gerontology. 2017. PMID: 28538216
Cited by
-
Resolving Geroplasticity to the Balance of Rejuvenins and Geriatrins.Aging Dis. 2022 Dec 1;13(6):1664-1714. doi: 10.14336/AD.2022.0414. eCollection 2022 Dec 1. Aging Dis. 2022. PMID: 36465174 Free PMC article.
-
The long and winding road of reprogramming-induced rejuvenation.Nat Commun. 2024 Mar 2;15(1):1941. doi: 10.1038/s41467-024-46020-5. Nat Commun. 2024. PMID: 38431638 Free PMC article. Review.
-
Rejuvenated Autologous Adult Stem Cells: Emerging Front Runners in the Fight Against Aging and Associated Diseases.Cells. 2025 Jul 25;14(15):1153. doi: 10.3390/cells14151153. Cells. 2025. PMID: 40801586 Free PMC article. Review.
-
A multiomic atlas of the aging hippocampus reveals molecular changes in response to environmental enrichment.Nat Commun. 2024 Jul 16;15(1):5829. doi: 10.1038/s41467-024-49608-z. Nat Commun. 2024. PMID: 39013876 Free PMC article.
-
Epigenetic rejuvenation: a journey backwards towards an epigenomic ground state.Epigenomics. 2025 Jan;17(1):1-3. doi: 10.1080/17501911.2024.2432851. Epub 2024 Nov 25. Epigenomics. 2025. PMID: 39584805 No abstract available.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
