

Neuronal and glial 3D chromatin architecture informs the cellular etiology of brain disorders
- PMID: 34172755
- PMCID: PMC8233376
- DOI: 10.1038/s41467-021-24243-0
Neuronal and glial 3D chromatin architecture informs the cellular etiology of brain disorders
Abstract
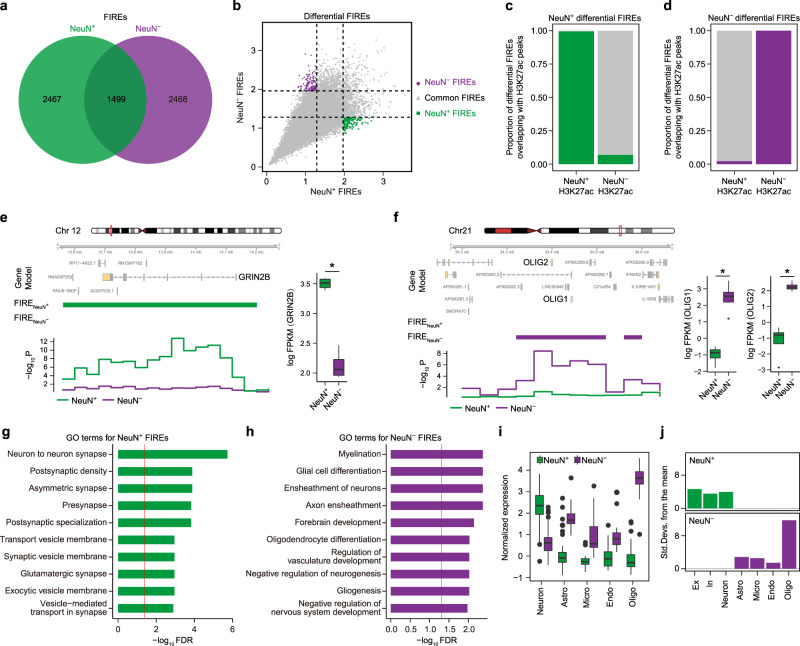
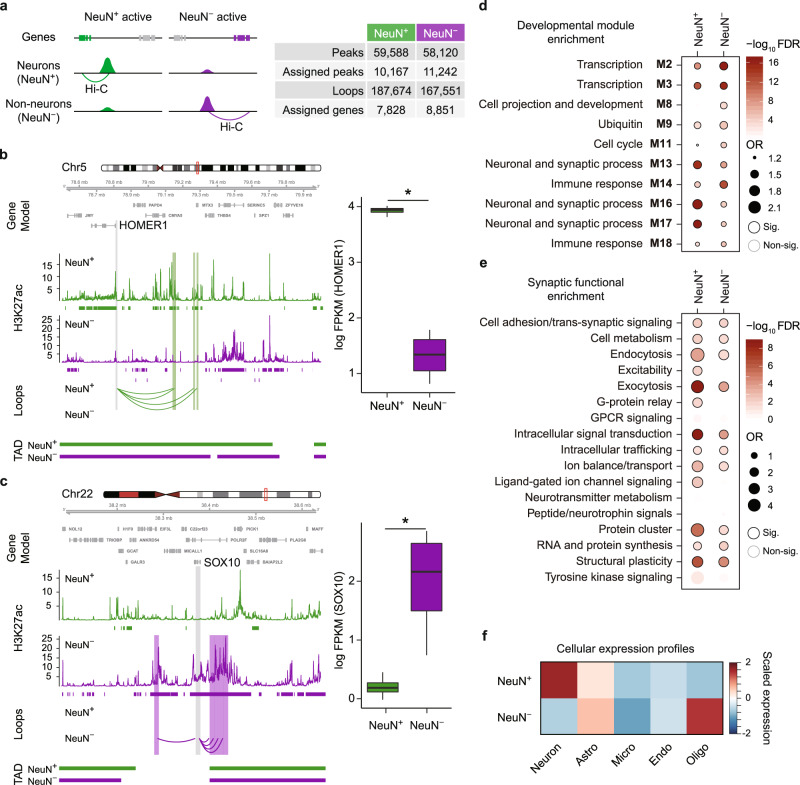
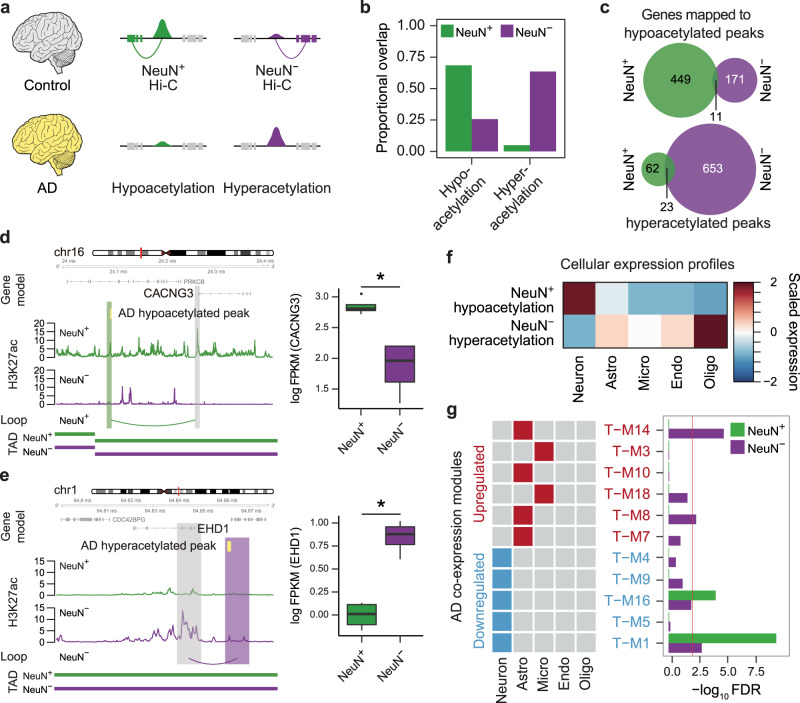
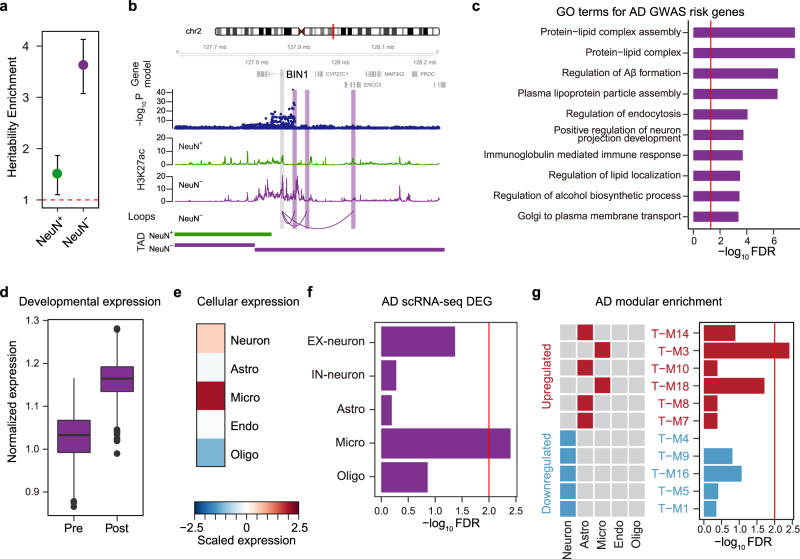
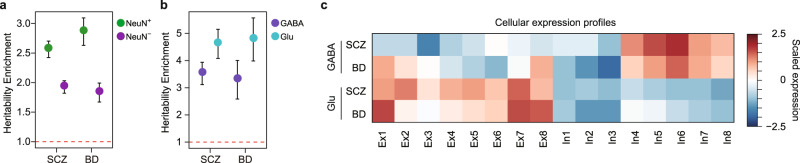
Cellular heterogeneity in the human brain obscures the identification of robust cellular regulatory networks, which is necessary to understand the function of non-coding elements and the impact of non-coding genetic variation. Here we integrate genome-wide chromosome conformation data from purified neurons and glia with transcriptomic and enhancer profiles, to characterize the gene regulatory landscape of two major cell classes in the human brain. We then leverage cell-type-specific regulatory landscapes to gain insight into the cellular etiology of several brain disorders. We find that Alzheimer's disease (AD)-associated epigenetic dysregulation is linked to neurons and oligodendrocytes, whereas genetic risk factors for AD highlighted microglia, suggesting that different cell types may contribute to disease risk, via different mechanisms. Moreover, integration of glutamatergic and GABAergic regulatory maps with genetic risk factors for schizophrenia (SCZ) and bipolar disorder (BD) identifies shared (parvalbumin-expressing interneurons) and distinct cellular etiologies (upper layer neurons for BD, and deeper layer projection neurons for SCZ). Collectively, these findings shed new light on cell-type-specific gene regulatory networks in brain disorders.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Rada-Iglesias A. Genetic variation within transcriptional regulatory elements and its implications for human disease. Biol. Chem. 2014;395:1453–1460. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 MH094714/MH/NIMH NIH HHS/United States
- I01 BX005585/BX/BLRD VA/United States
- P50 MH106438/MH/NIMH NIH HHS/United States
- R01 MH100027/MH/NIMH NIH HHS/United States
- R01 MH110927/MH/NIMH NIH HHS/United States
- U01 MH103392/MH/NIMH NIH HHS/United States
- U01 MH122590/MH/NIMH NIH HHS/United States
- R21 DA051921/DA/NIDA NIH HHS/United States
- U01 MH103339/MH/NIMH NIH HHS/United States
- DP2 MH122403/MH/NIMH NIH HHS/United States
- P50 MH096890/MH/NIMH NIH HHS/United States
- R01 MH110926/MH/NIMH NIH HHS/United States
- I01 BX002876/BX/BLRD VA/United States
- R21 MH103877/MH/NIMH NIH HHS/United States
- R00 MH113823/MH/NIMH NIH HHS/United States
- P50 HD103573/HD/NICHD NIH HHS/United States
- R01 AG066871/AG/NIA NIH HHS/United States
- R01 MH121521/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
