

Hepatic resistance to cold ferroptosis in a mammalian hibernator Syrian hamster depends on effective storage of diet-derived α-tocopherol
- PMID: 34172811
- PMCID: PMC8233303
- DOI: 10.1038/s42003-021-02297-6
Hepatic resistance to cold ferroptosis in a mammalian hibernator Syrian hamster depends on effective storage of diet-derived α-tocopherol
Abstract
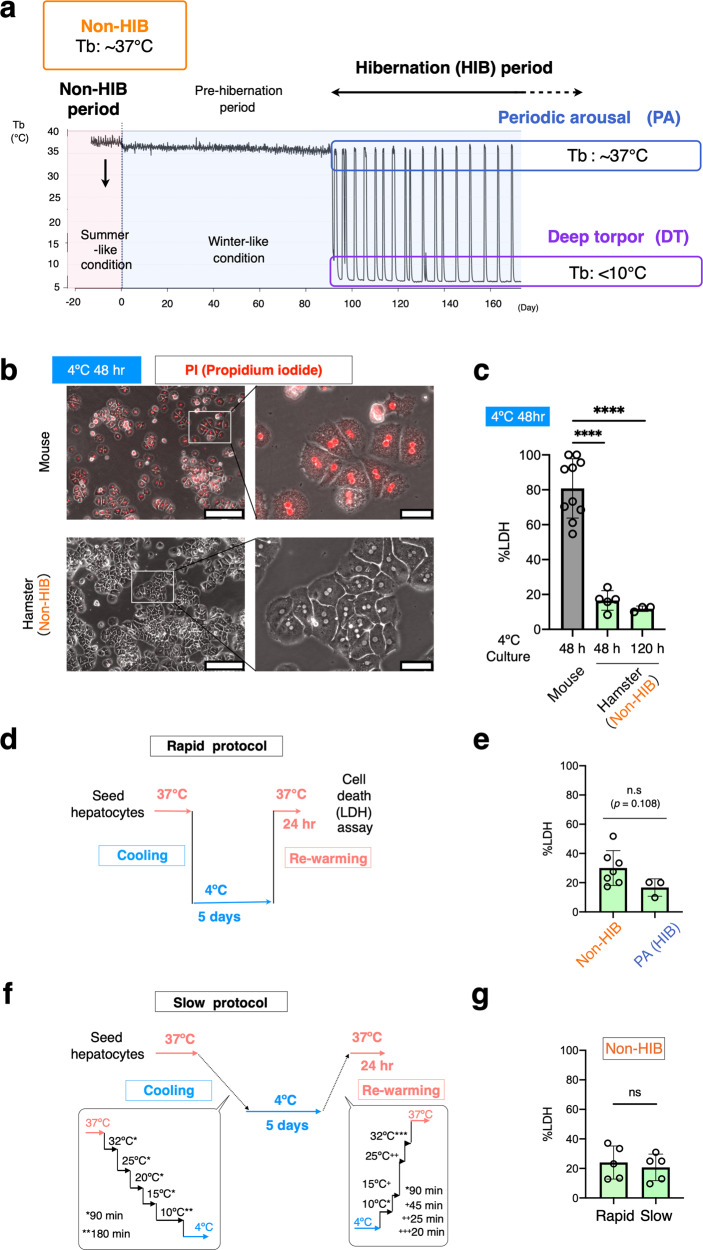
Mammalian hibernators endure severe and prolonged hypothermia that is lethal to non-hibernators, including humans and mice. The mechanisms responsible for the cold resistance remain poorly understood. Here, we found that hepatocytes from a mammalian hibernator, the Syrian hamster, exhibited remarkable resistance to prolonged cold culture, whereas murine hepatocytes underwent cold-induced cell death that fulfills the hallmarks of ferroptosis such as necrotic morphology, lipid peroxidation and prevention by an iron chelator. Unexpectedly, hepatocytes from Syrian hamsters exerted resistance to cold- and drug-induced ferroptosis in a diet-dependent manner, with the aid of their superior ability to retain dietary α-tocopherol (αT), a vitamin E analog, in the liver and blood compared with those of mice. The liver phospholipid composition is less susceptible to peroxidation in Syrian hamsters than in mice. Altogether, the cold resistance of the hibernator's liver is established by the ability to utilize αT effectively to prevent lipid peroxidation and ferroptosis.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Andrews, M. T. Molecular interactions underpinning the phenotype of hibernation in mammals. J. Exp. Biol. 222, 10.1242/jeb.160606 (2019). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
