

From synapse to network: models of information storage and retrieval in neural circuits
- PMID: 34175521
- PMCID: PMC8688181
- DOI: 10.1016/j.conb.2021.05.005
From synapse to network: models of information storage and retrieval in neural circuits
Abstract
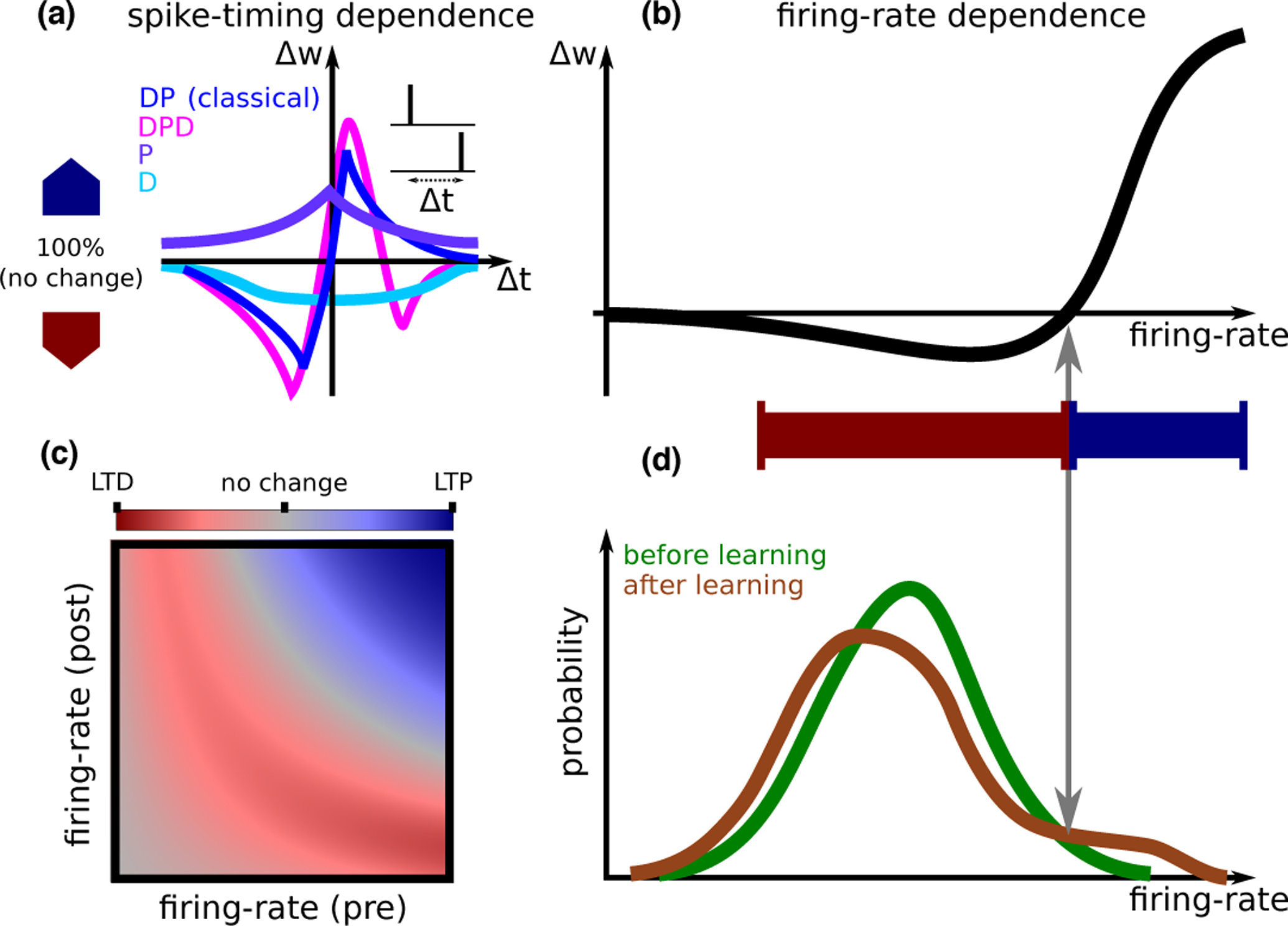
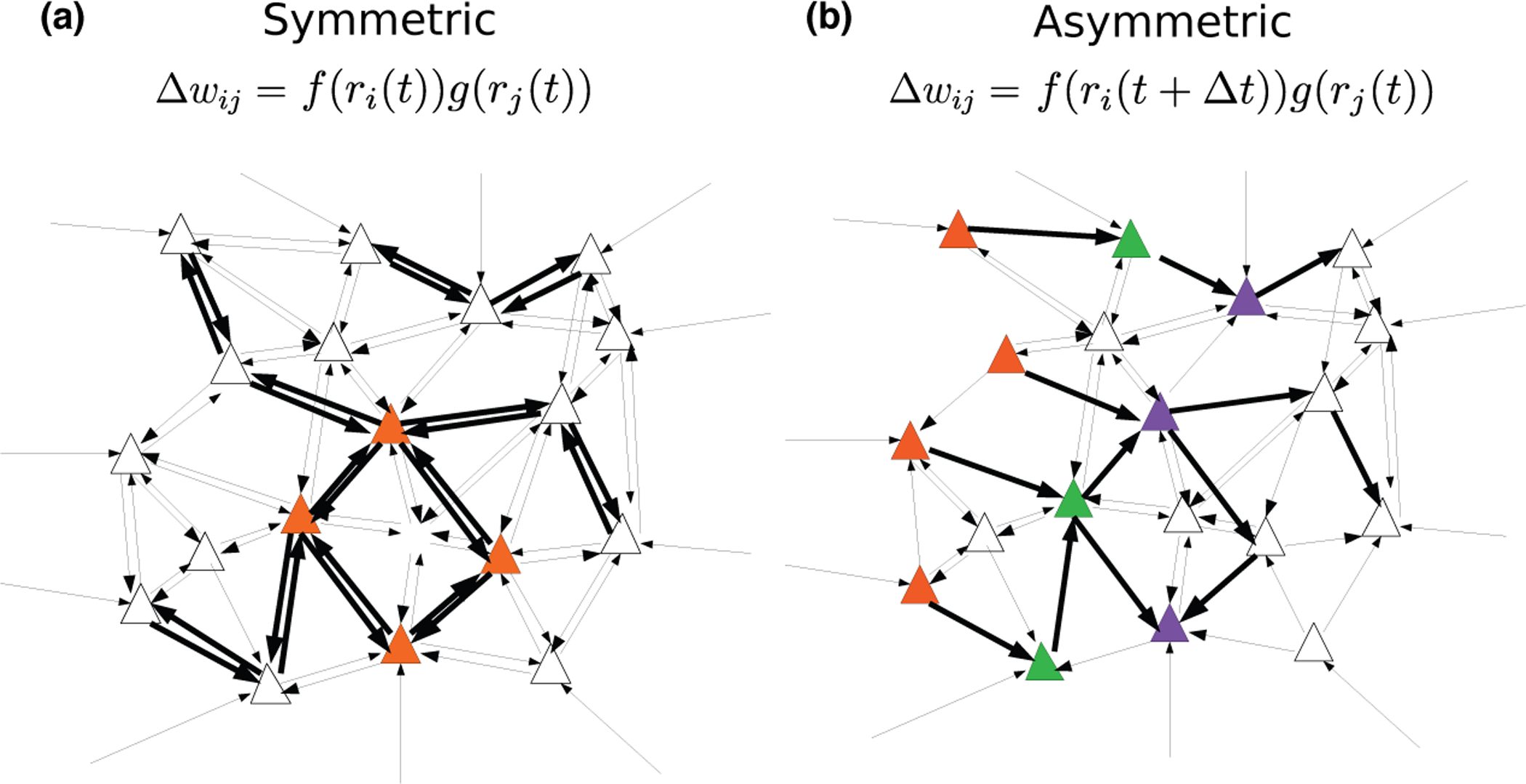
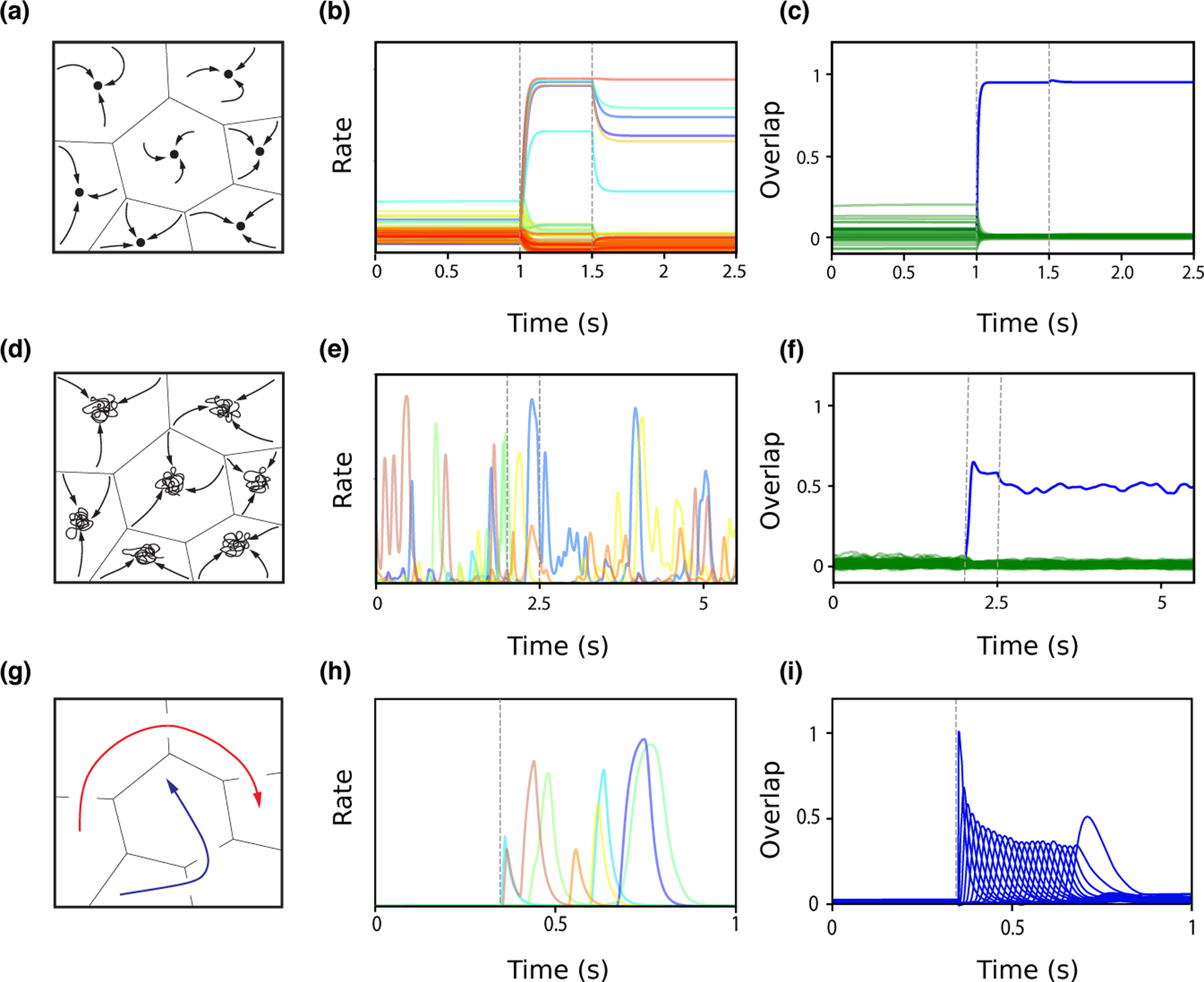
The mechanisms of information storage and retrieval in brain circuits are still the subject of debate. It is widely believed that information is stored at least in part through changes in synaptic connectivity in networks that encode this information and that these changes lead in turn to modifications of network dynamics, such that the stored information can be retrieved at a later time. Here, we review recent progress in deriving synaptic plasticity rules from experimental data and in understanding how plasticity rules affect the dynamics of recurrent networks. We show that the dynamics generated by such networks exhibit a large degree of diversity, depending on parameters, similar to experimental observations in vivo during delayed response tasks.
Copyright © 2021 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest statement Nothing declared.
Figures
References
-
- Markram H, Lubke J, Frotscher M, and Sakmann B (1997). Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 275, 213–215. - PubMed
-
- Inglebert Y, Aljade J, Brunel N, and Debanne D (2020). Synaptic plasticity rules with physiological calcium levels. Proc Natl Acad Sci U S A 117, 33639–33648. - PMC - PubMed
-
•STDP experiments at physiological calcium concentrations show no significant synaptic changes, while high firing rates and/or bursts restore plasticity. Experimental data is consistent with a calcium-based rule with non-linear summation between calcium transients due to pre and post-synaptic spikes.
