

α-Ketoglutarate Upregulates Collecting Duct (Pro)renin Receptor Expression, Tubular Angiotensin II Formation, and Na+ Reabsorption During High Glucose Conditions
- PMID: 34179130
- PMCID: PMC8220822
- DOI: 10.3389/fcvm.2021.644797
α-Ketoglutarate Upregulates Collecting Duct (Pro)renin Receptor Expression, Tubular Angiotensin II Formation, and Na+ Reabsorption During High Glucose Conditions
Abstract
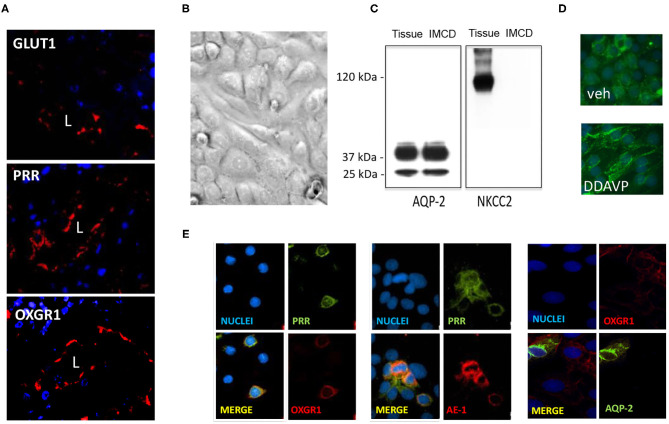
Diabetes mellitus (DM) causes high glucose (HG) levels in the plasma and urine. The (pro)renin receptor (PRR) is a key regulator of renal Na+ handling. PRR is expressed in intercalated (IC) cells of the collecting duct (CD) and binds renin to promote angiotensin (Ang) II formation, thereby contributing to Na+ reabsorption. In DM, the Kreb's cycle is in a state of suppression in most tissues. However, in the CD, expression of glucose transporters is augmented, boosting the Kreb's cycle and consequently causing α-ketoglutarate (αKG) accumulation. The αKG receptor 1 (OXGR1) is a Gq-coupled receptor expressed on the apical membrane of IC cells of the CD. We hypothesize that HG causes αKG secretion and activation of OXGR1, which increases PRR expression in CD cells. This effect then promotes intratubular AngII formation and Na+ reabsorption. To test this hypothesis, streptozotocin (STZ)-induced diabetic mice were treated with or without montelukast (ML), an OXGR1 antagonist, for 6 days. STZ mice had higher urinary αKG and PRR expression along with augmented urinary AngII levels and Na+ retention. Treatment with ML prevented all these effects. Similarly, primary cultured inner medullary CD cells treated with HG showed increased PRR expression, while OXGR1 antagonist prevented this effect. αKG increases PRR expression, while treatments with ML, PKC inhibition, or intracellular Ca2+ depletion impair this effect. In silico analysis suggested that αKG binds to mouse OXGR1. These results indicate that HG conditions promote increased levels of intratubular αKG and OXGR1-dependent PRR upregulation, which impact AngII formation and Na+ reabsorption.
Keywords: Kreb's cycle; angiotensin; collecting duct; diabetes; prorenin receptor.
Copyright © 2021 Guerrero, Visniauskas, Cárdenas, Figueroa, Vivanco, Salinas-Parra, Araos, Nguyen, Kassan, Amador, Prieto and Gonzalez.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous