

Neural specification, targeting, and circuit formation during visual system assembly
- PMID: 34183440
- PMCID: PMC8285955
- DOI: 10.1073/pnas.2101823118
Neural specification, targeting, and circuit formation during visual system assembly
Abstract
Like other sensory systems, the visual system is topographically organized: Its sensory neurons, the photoreceptors, and their targets maintain point-to-point correspondence in physical space, forming a retinotopic map. The iterative wiring of circuits in the visual system conveniently facilitates the study of its development. Over the past few decades, experiments in Drosophila have shed light on the principles that guide the specification and connectivity of visual system neurons. In this review, we describe the main findings unearthed by the study of the Drosophila visual system and compare them with similar events in mammals. We focus on how temporal and spatial patterning generates diverse cell types, how guidance molecules distribute the axons and dendrites of neurons within the correct target regions, how vertebrates and invertebrates generate their retinotopic map, and the molecules and mechanisms required for neuronal migration. We suggest that basic principles used to wire the fly visual system are broadly applicable to other systems and highlight its importance as a model to study nervous system development.
Keywords: Drosophila; neural development; patterning; retina; visual system.
Conflict of interest statement
The authors declare no competing interest.
Figures
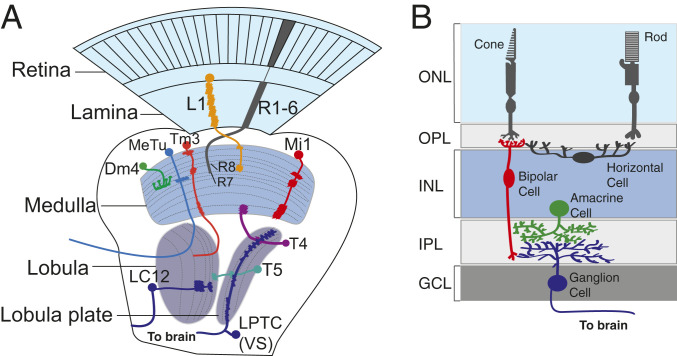
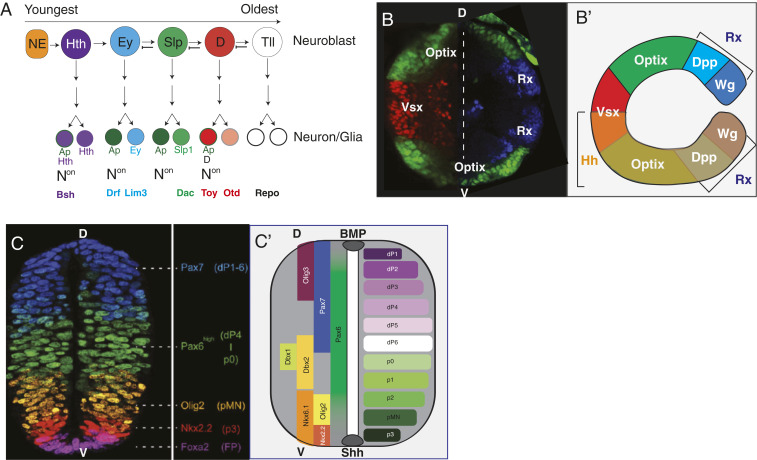
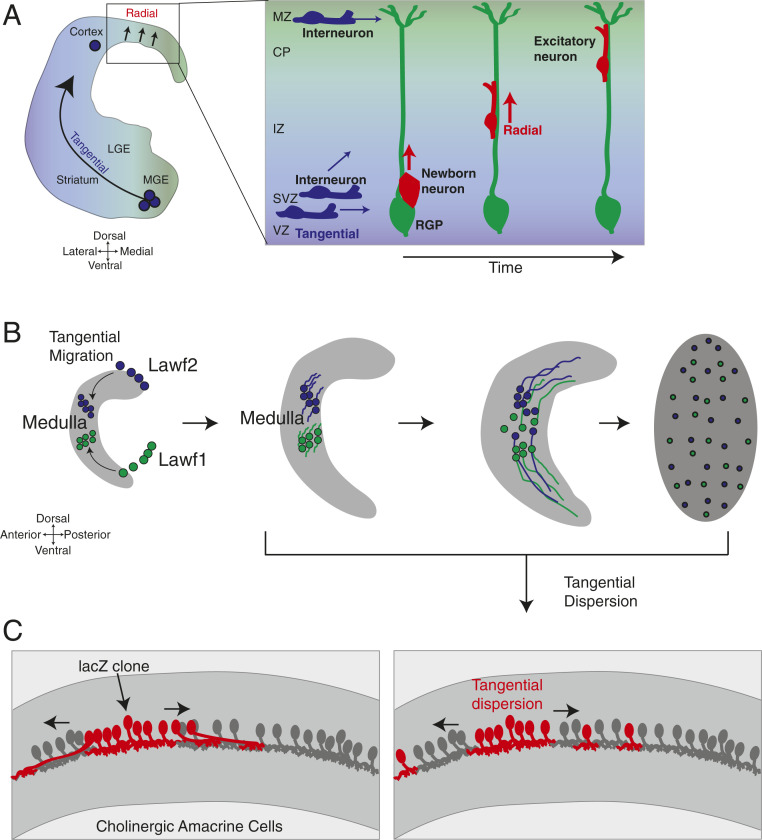
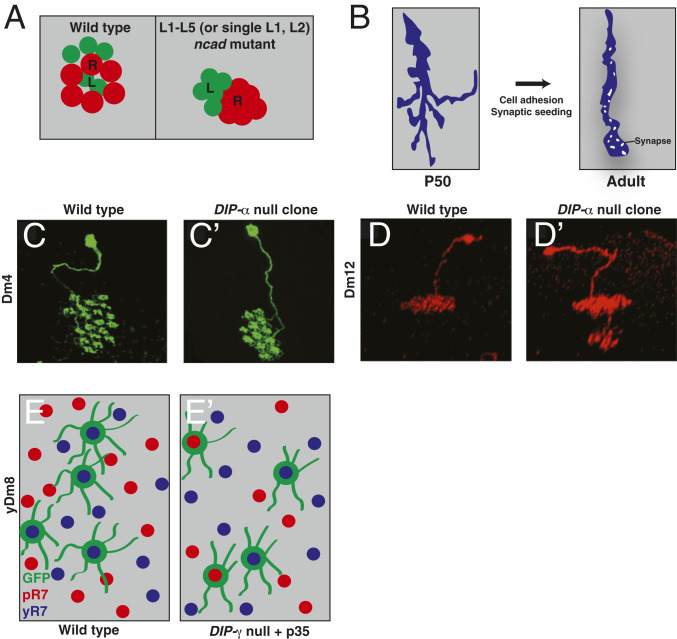
References
-
- Luo L., Flanagan J. G., Development of continuous and discrete neural maps. Neuron 56, 284–300 (2007). - PubMed
-
- Fischbach K.-F., Dittrich A. P. M., The optic lobe of Drosophila melanogaster. I. A Golgi analysis of wild-type structure. Cell Tissue Res. 258, 441–475 (1989).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
