

Ki-67 gene expression
- PMID: 34183782
- PMCID: PMC8629999
- DOI: 10.1038/s41418-021-00823-x
Ki-67 gene expression
Abstract
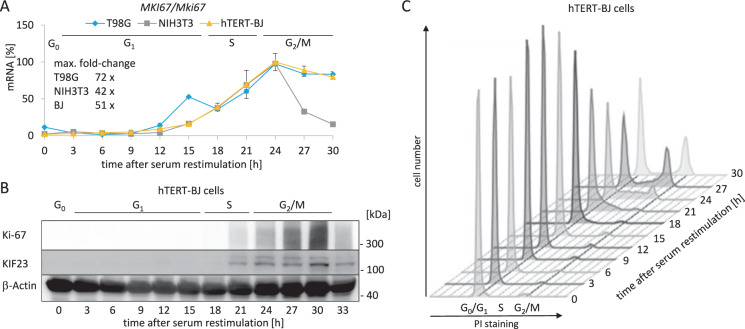
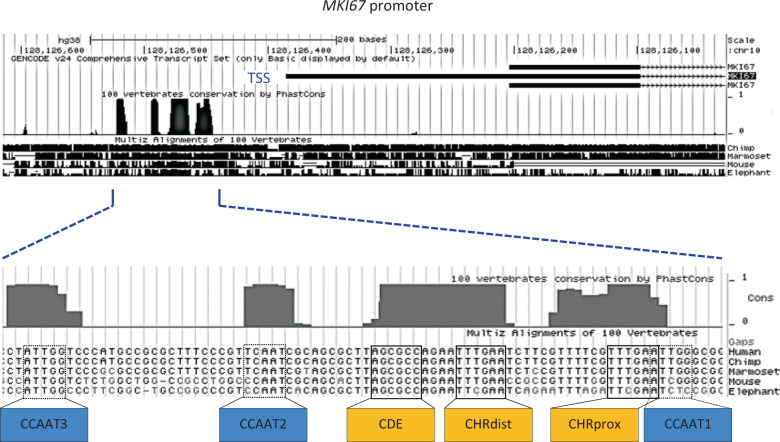
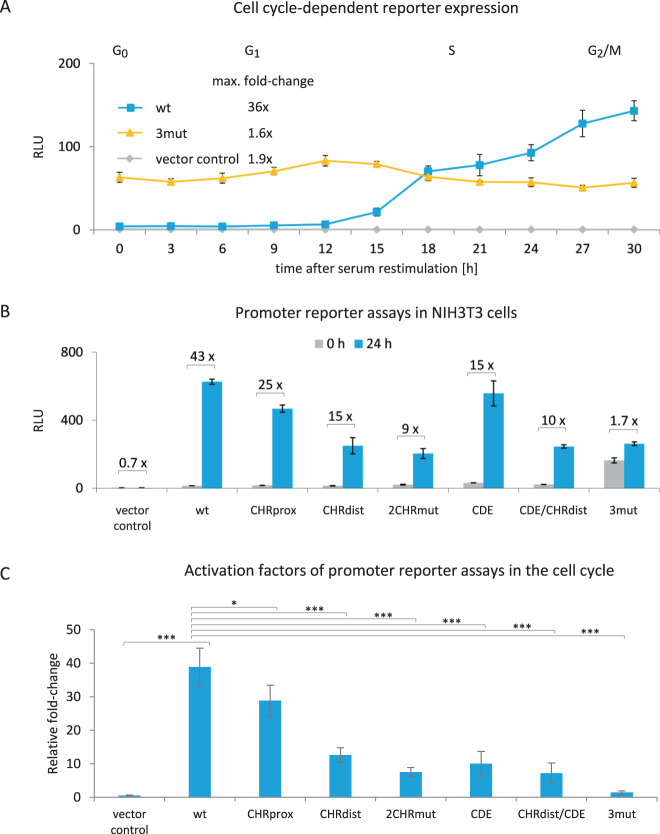
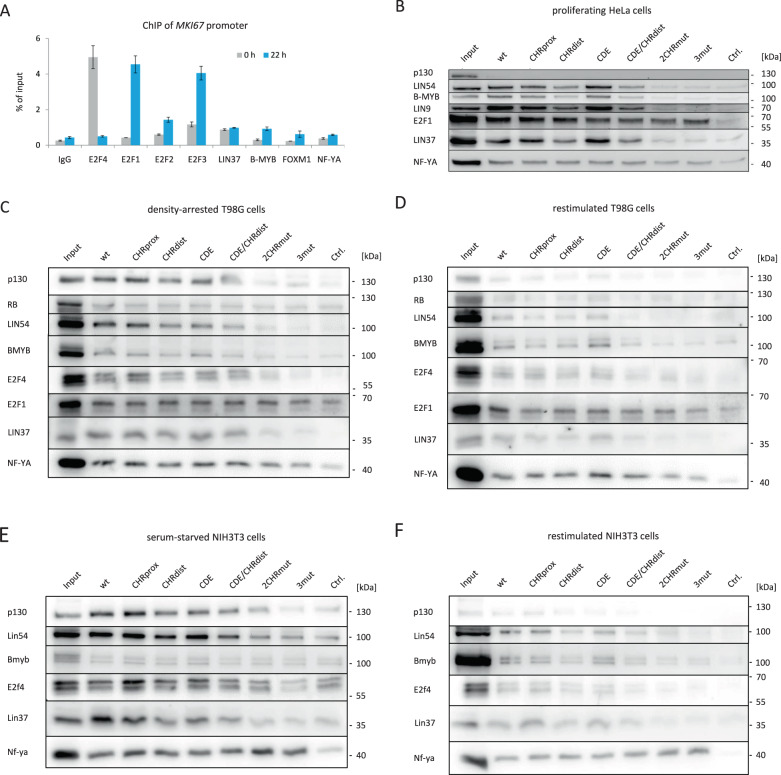
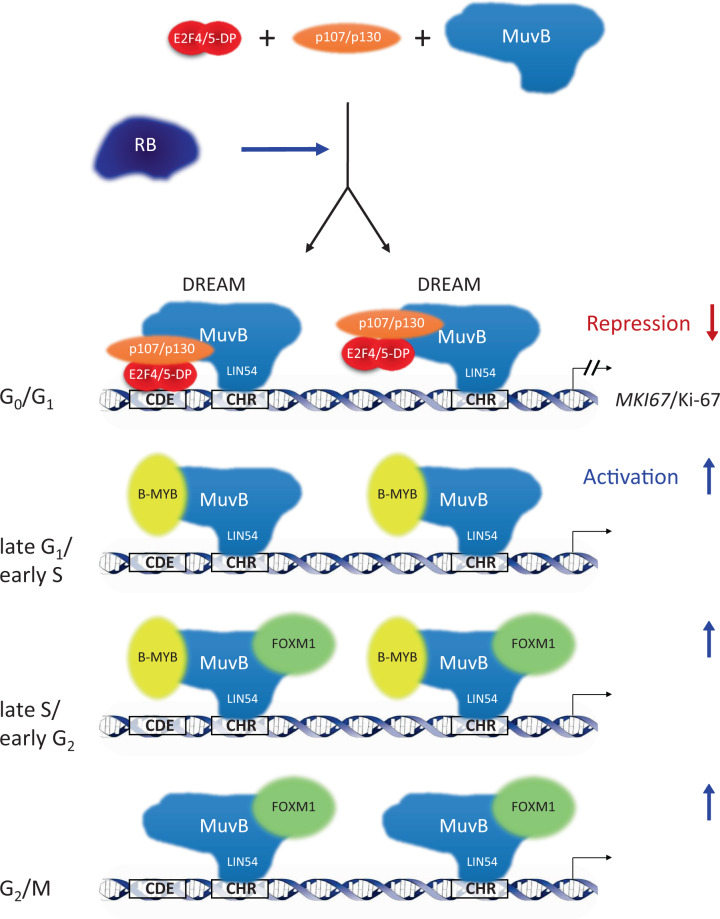
Ki-67 serves as a prominent cancer marker. We describe how expression of the MKI67 gene coding for Ki-67 is controlled during the cell cycle. MKI67 mRNA and Ki-67 protein are maximally expressed in G2 phase and mitosis. Expression is dependent on two CHR elements and one CDE site in the MKI67 promoter. DREAM transcriptional repressor complexes bind to both CHR sites and downregulate the expression in G0/G1 cells. Upregulation of MKI67 transcription coincides with binding of B-MYB-MuvB and FOXM1-MuvB complexes from S phase into G2/M. Importantly, binding of B-MYB to the two CHR elements correlates with loss of CHR-dependent MKI67 promoter activation in B-MYB-knockdown experiments. In knockout cell models, we find that DREAM/MuvB-dependent transcriptional control cooperates with the RB Retinoblastoma tumor suppressor. Furthermore, the p53 tumor suppressor indirectly downregulates transcription of the MKI67 gene. This repression by p53 requires p21/CDKN1A. These results are consistent with a model in which DREAM, B-MYB-MuvB, and FOXM1-MuvB together with RB cooperate in cell cycle-dependent transcription and in transcriptional repression following p53 activation. In conclusion, we present mechanisms how MKI67 gene expression followed by Ki-67 protein synthesis is controlled during the cell cycle and upon induction of DNA damage, as well as upon p53 activation.
© 2021. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
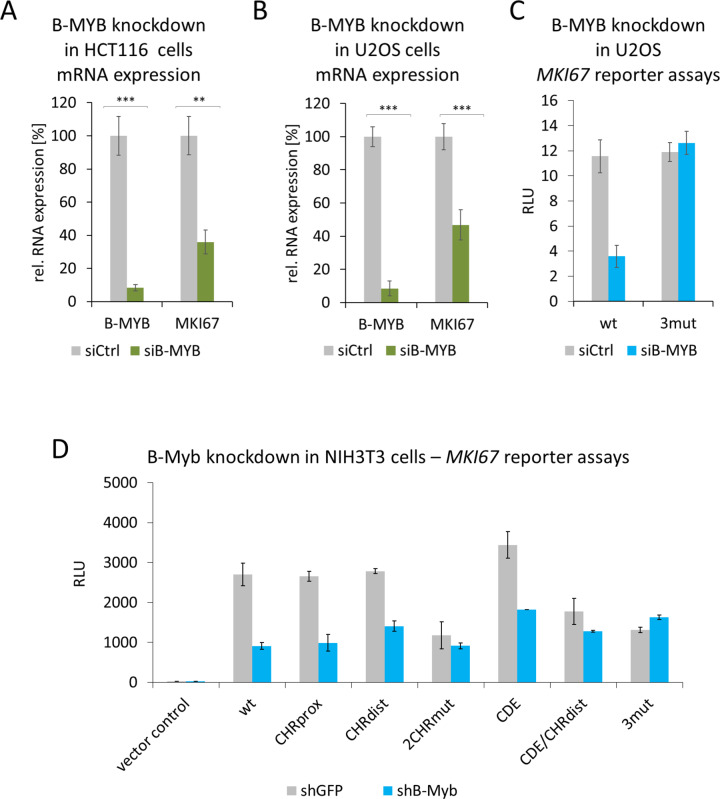
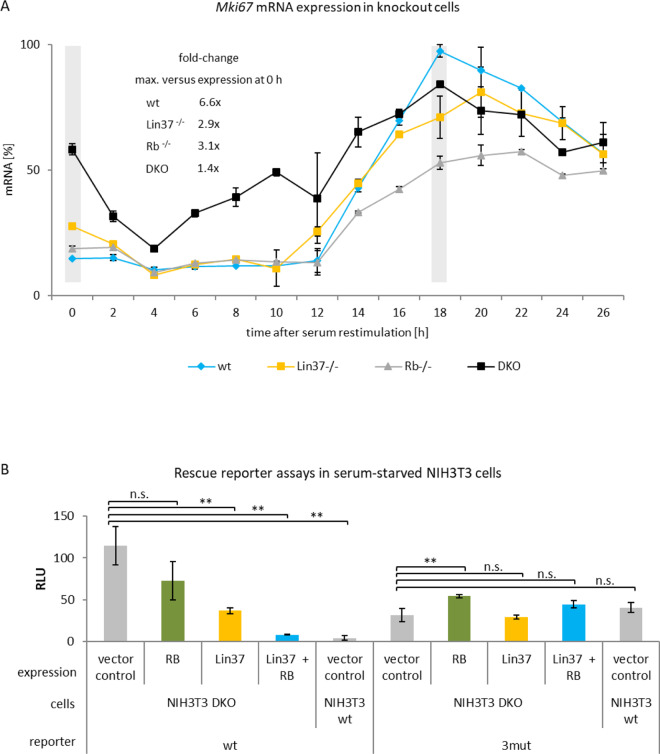
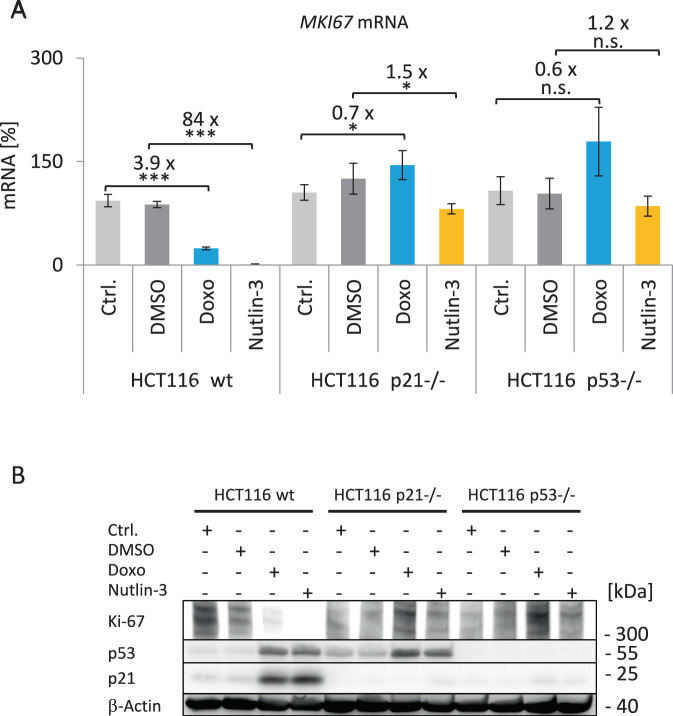
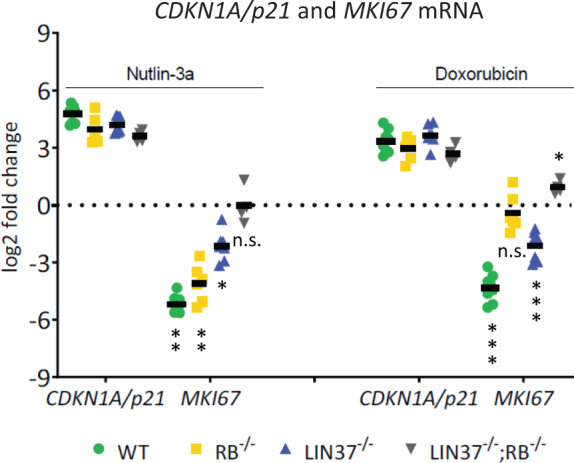
References
-
- Gerdes J, Schwab U, Lemke H, Stein H. Production of a mouse monoclonal antibody reactive with a human nuclear antigen associated with cell proliferation. Int J Cancer. 1983;31:13–20. - PubMed
-
- Scholzen T, Gerdes J. The Ki-67 protein: from the known and the unknown. J Cell Physiol. 2000;182:311–22. - PubMed
-
- Gerdes J, Lemke H, Baisch H, Wacker HH, Schwab U, Stein H. Cell cycle analysis of a cell proliferation-associated human nuclear antigen defined by the monoclonal antibody Ki-67. J Immunol. 1984;133:1710–5. - PubMed
-
- Dowsett M, Dunbier AK. Emerging biomarkers and new understanding of traditional markers in personalized therapy for breast cancer. Clin Cancer Res. 2008;14:8019–26. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
