

Self-activation of Vγ9Vδ2 T cells by exogenous phosphoantigens involves TCR and butyrophilins
- PMID: 34183807
- PMCID: PMC8237548
- DOI: 10.1038/s41423-021-00720-w
Self-activation of Vγ9Vδ2 T cells by exogenous phosphoantigens involves TCR and butyrophilins
Abstract
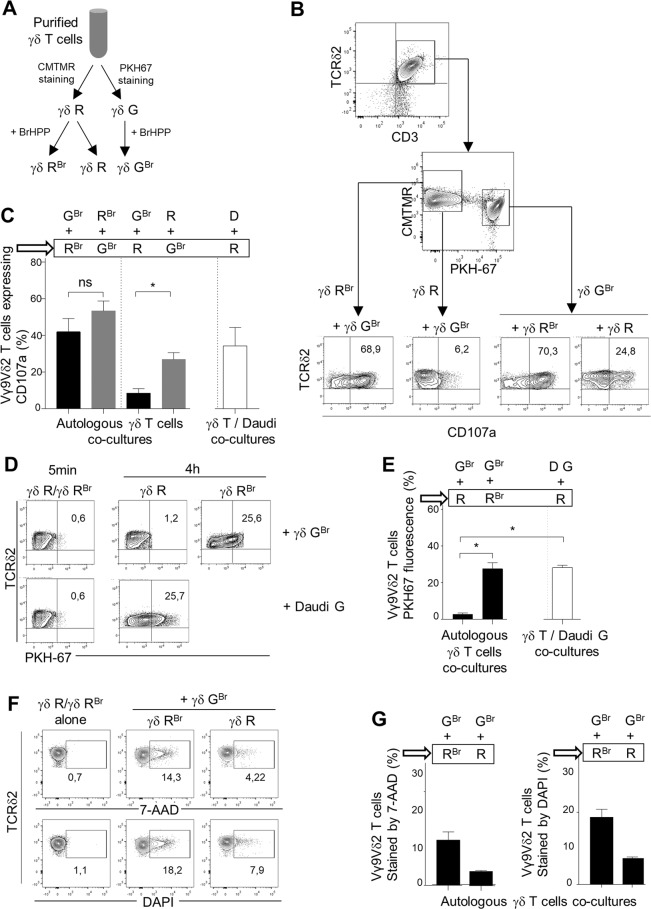
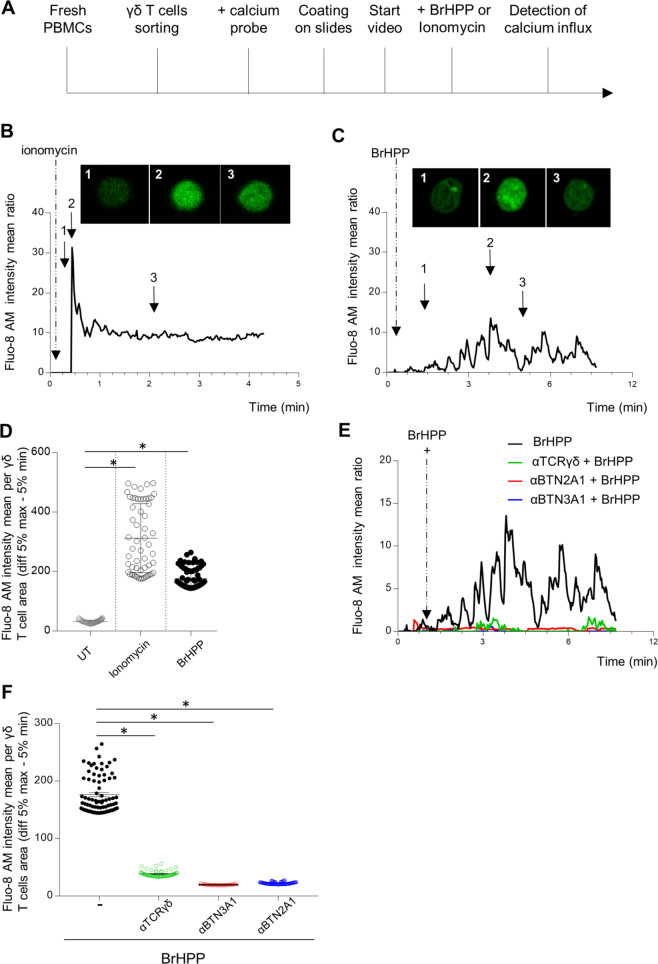
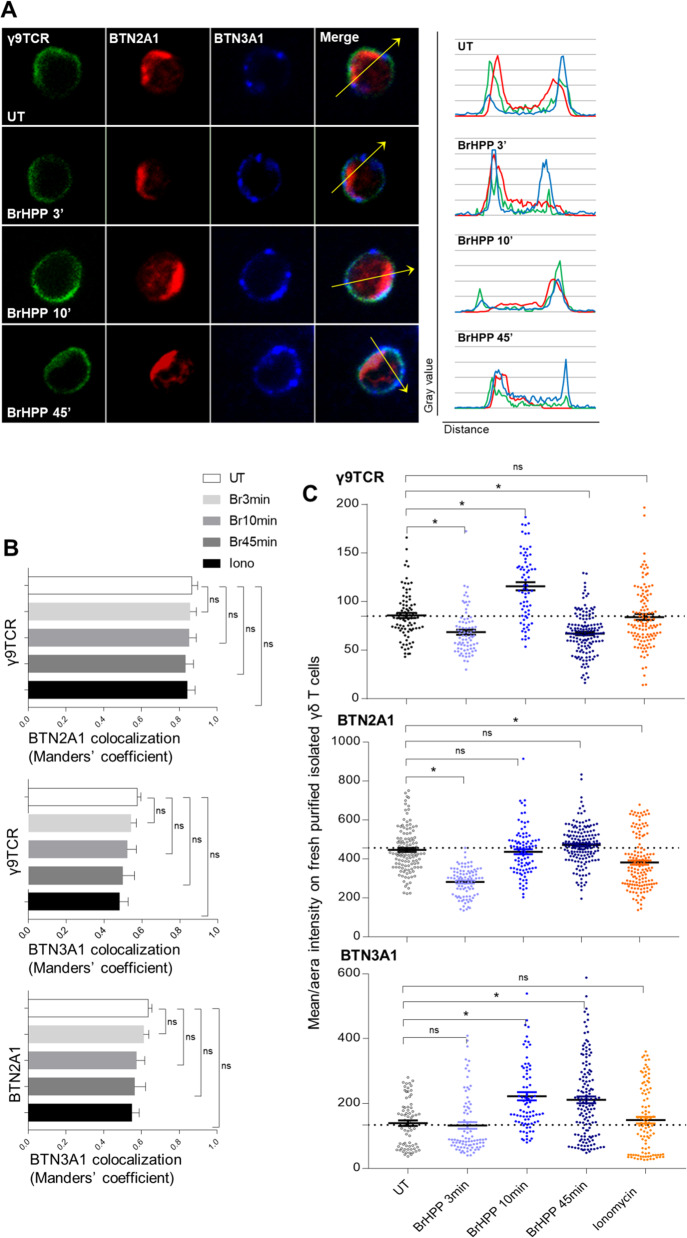
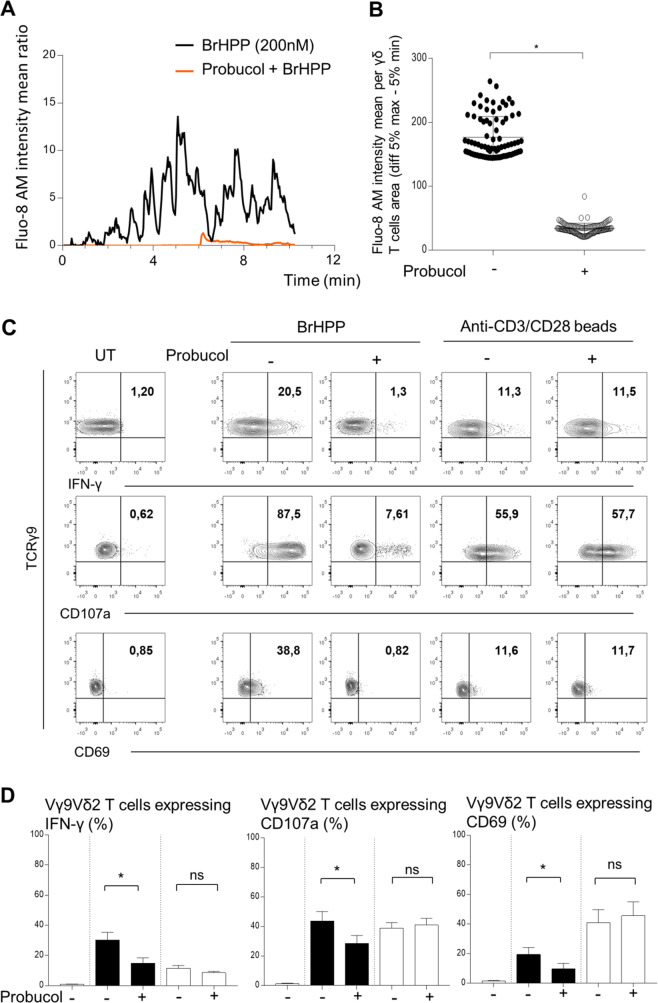
The high cytotoxic activity of Vγ9Vδ2 T lymphocytes against tumor cells makes them useful candidates in anticancer therapies. However, the molecular mechanism of their activation by phosphoantigens (PAgs) is not completely known. Many studies have depicted the mechanism of Vγ9Vδ2 T-cell activation by PAg-sensed accessory cells, such as immune presenting cells or tumor cells. In this study, we demonstrated that pure resting Vγ9Vδ2 T lymphocytes can self-activate through exogenous PAgs, involving their TCR and the butyrophilins BTN3A1 and BTN2A1. This is the first time that these three molecules, concurrently expressed at the plasma membrane of Vγ9Vδ2 T cells, have been shown to be involved together on the same and unique T cell during PAg activation. Moreover, the use of probucol to stimulate the inhibition of this self-activation prompted us to propose that ABCA-1 could be implicated in the transfer of exogenous PAgs inside Vγ9Vδ2 T cells before activating them through membrane clusters formed by γ9TCR, BTN3A1 and BTN2A1. The self-activation of Vγ9Vδ2 T cells, which leads to self-killing, can therefore participate in the failure of γδ T cell-based therapies with exogenous PAgs and should be taken into account.
Keywords: Butyrophilins; Phosphoantigen; T-cell receptor; Vγ9Vδ2 T cells.
© 2021. The Author(s), under exclusive licence to CSI and USTC.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Choudhary A, Davodeau F, Moreau A, Peyrat MA, Bonneville M, Jotereau F. Selective lysis of autologous tumor cells by recurrent gamma delta tumor-infiltrating lymphocytes from renal carcinoma. J Immunol. 1995;154:3932–40. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
