

Efficient C•G-to-G•C base editors developed using CRISPRi screens, target-library analysis, and machine learning
- PMID: 34183861
- PMCID: PMC8985520
- DOI: 10.1038/s41587-021-00938-z
Efficient C•G-to-G•C base editors developed using CRISPRi screens, target-library analysis, and machine learning
Erratum in
-
Author Correction: Efficient C•G-to-G•C base editors developed using CRISPRi screens, target-library analysis, and machine learning.Nat Biotechnol. 2023 Nov;41(11):1655. doi: 10.1038/s41587-023-02028-8. Nat Biotechnol. 2023. PMID: 37853259 No abstract available.
Abstract
Programmable C•G-to-G•C base editors (CGBEs) have broad scientific and therapeutic potential, but their editing outcomes have proved difficult to predict and their editing efficiency and product purity are often low. We describe a suite of engineered CGBEs paired with machine learning models to enable efficient, high-purity C•G-to-G•C base editing. We performed a CRISPR interference (CRISPRi) screen targeting DNA repair genes to identify factors that affect C•G-to-G•C editing outcomes and used these insights to develop CGBEs with diverse editing profiles. We characterized ten promising CGBEs on a library of 10,638 genomically integrated target sites in mammalian cells and trained machine learning models that accurately predict the purity and yield of editing outcomes (R = 0.90) using these data. These CGBEs enable correction to the wild-type coding sequence of 546 disease-related transversion single-nucleotide variants (SNVs) with >90% precision (mean 96%) and up to 70% efficiency (mean 14%). Computational prediction of optimal CGBE-single-guide RNA pairs enables high-purity transversion base editing at over fourfold more target sites than achieved using any single CGBE variant.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Figures
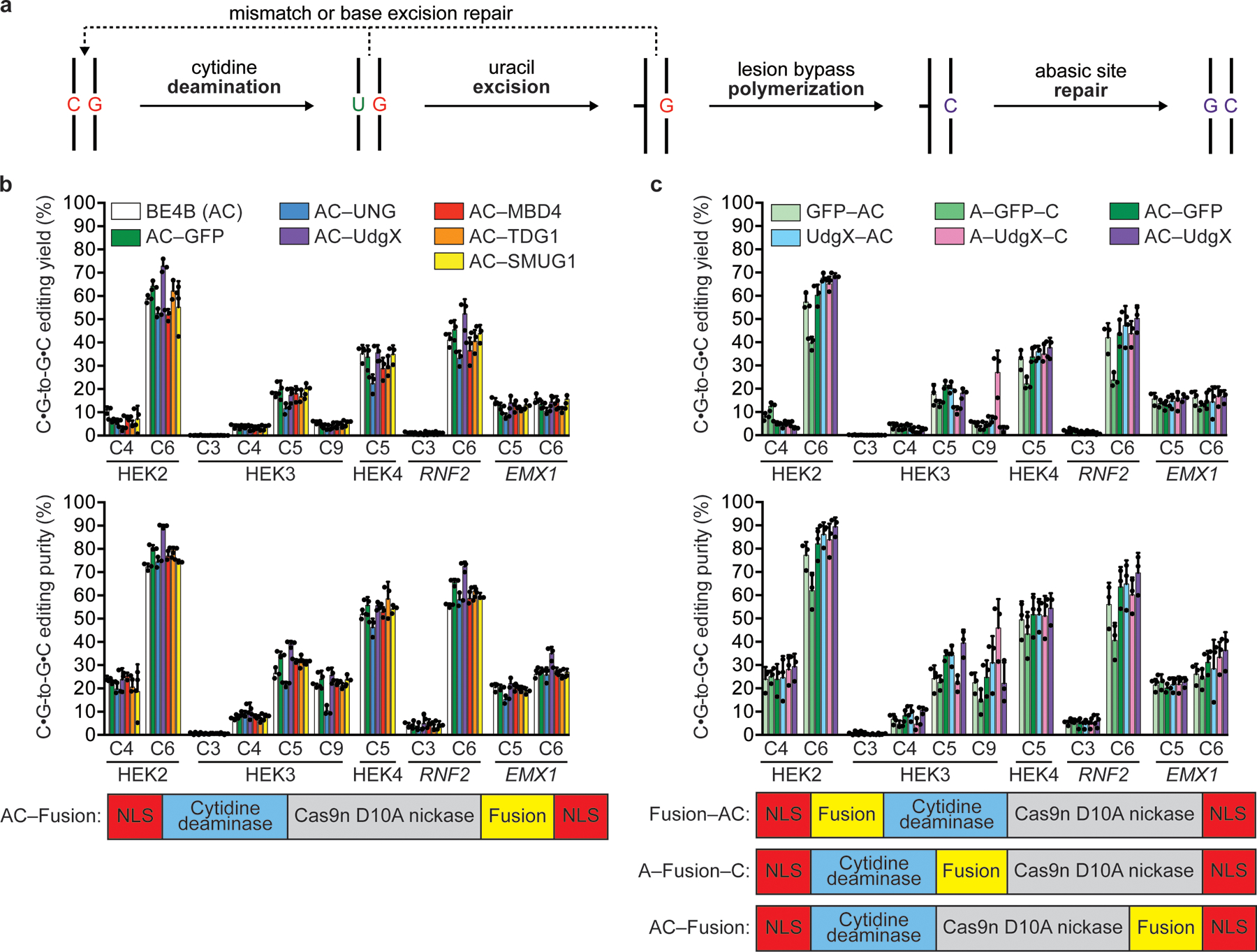
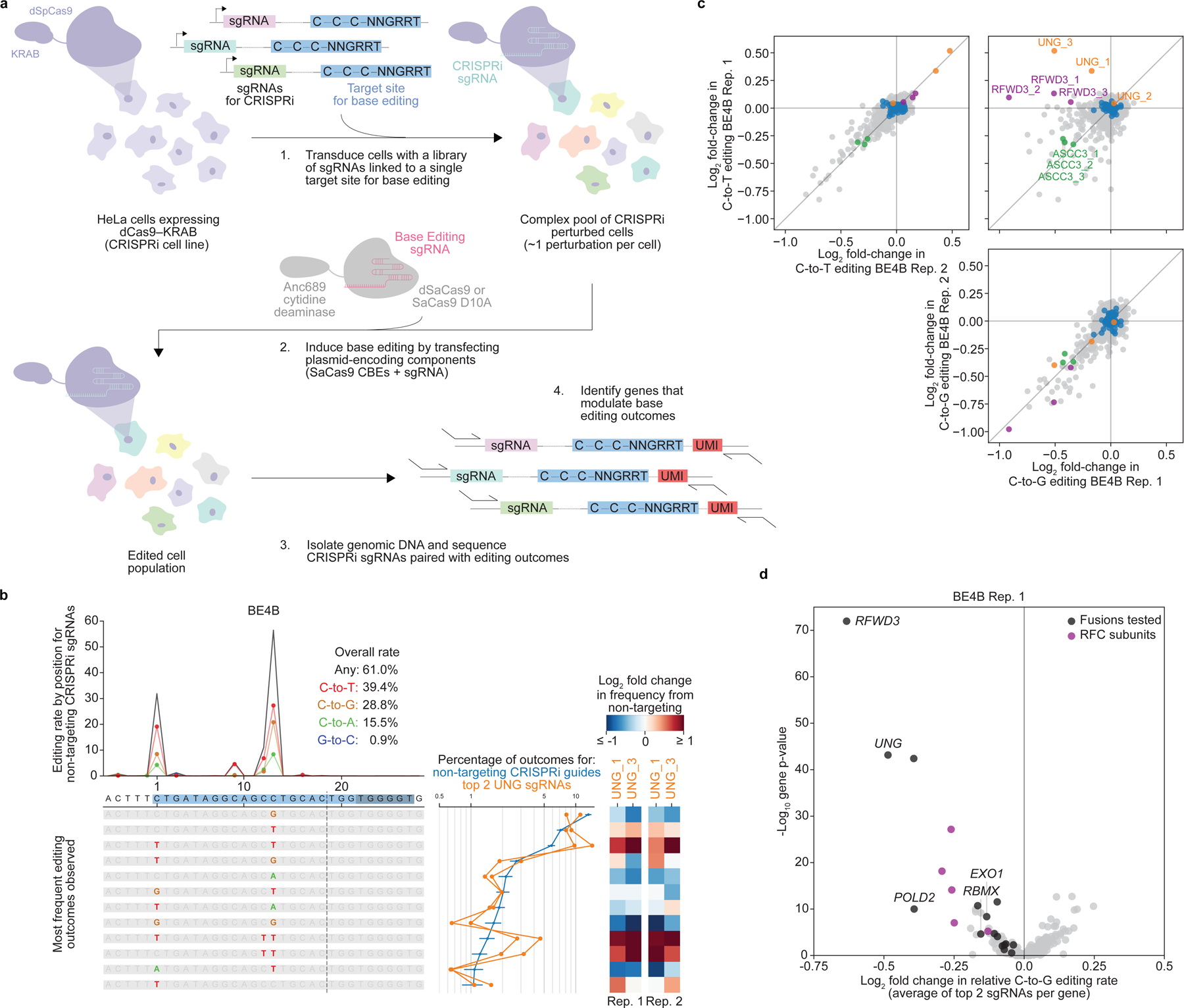
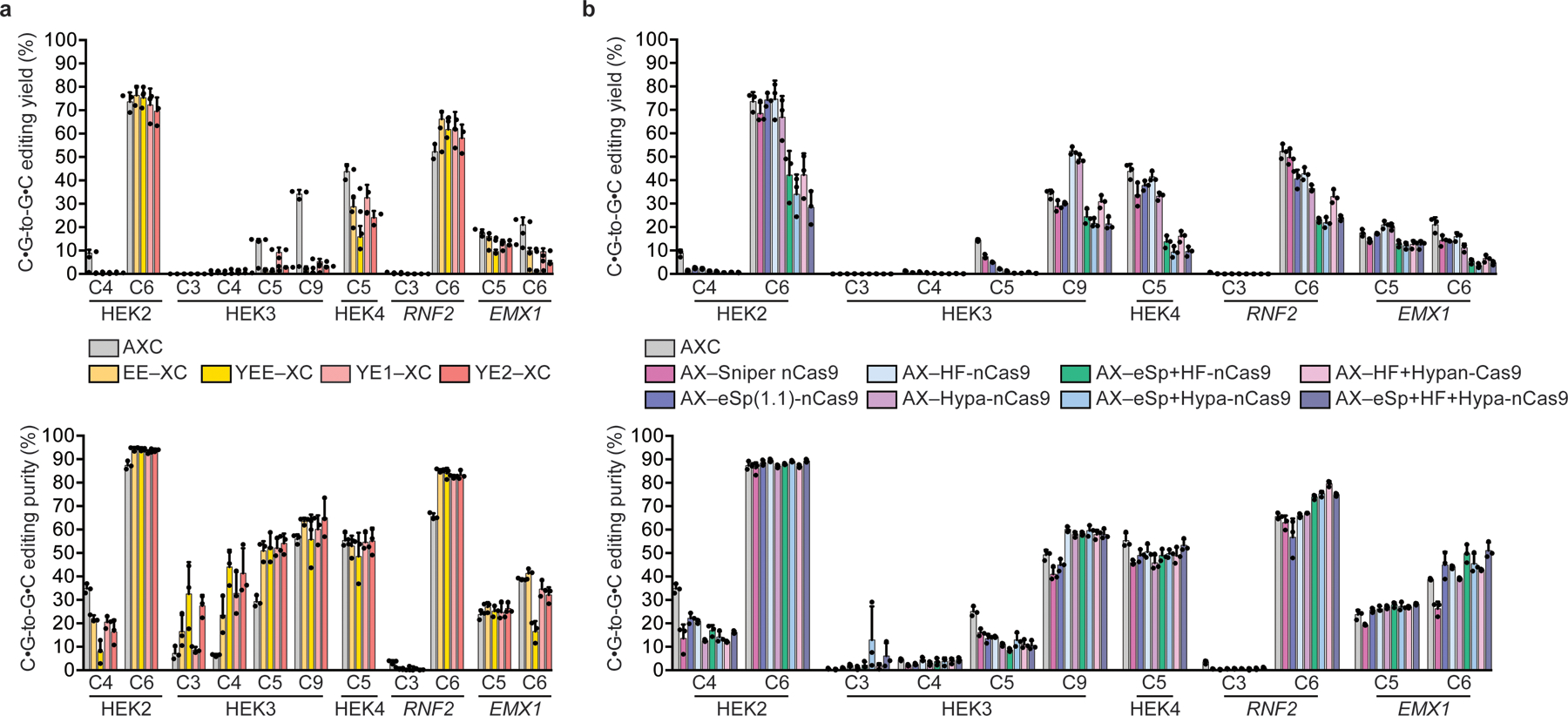
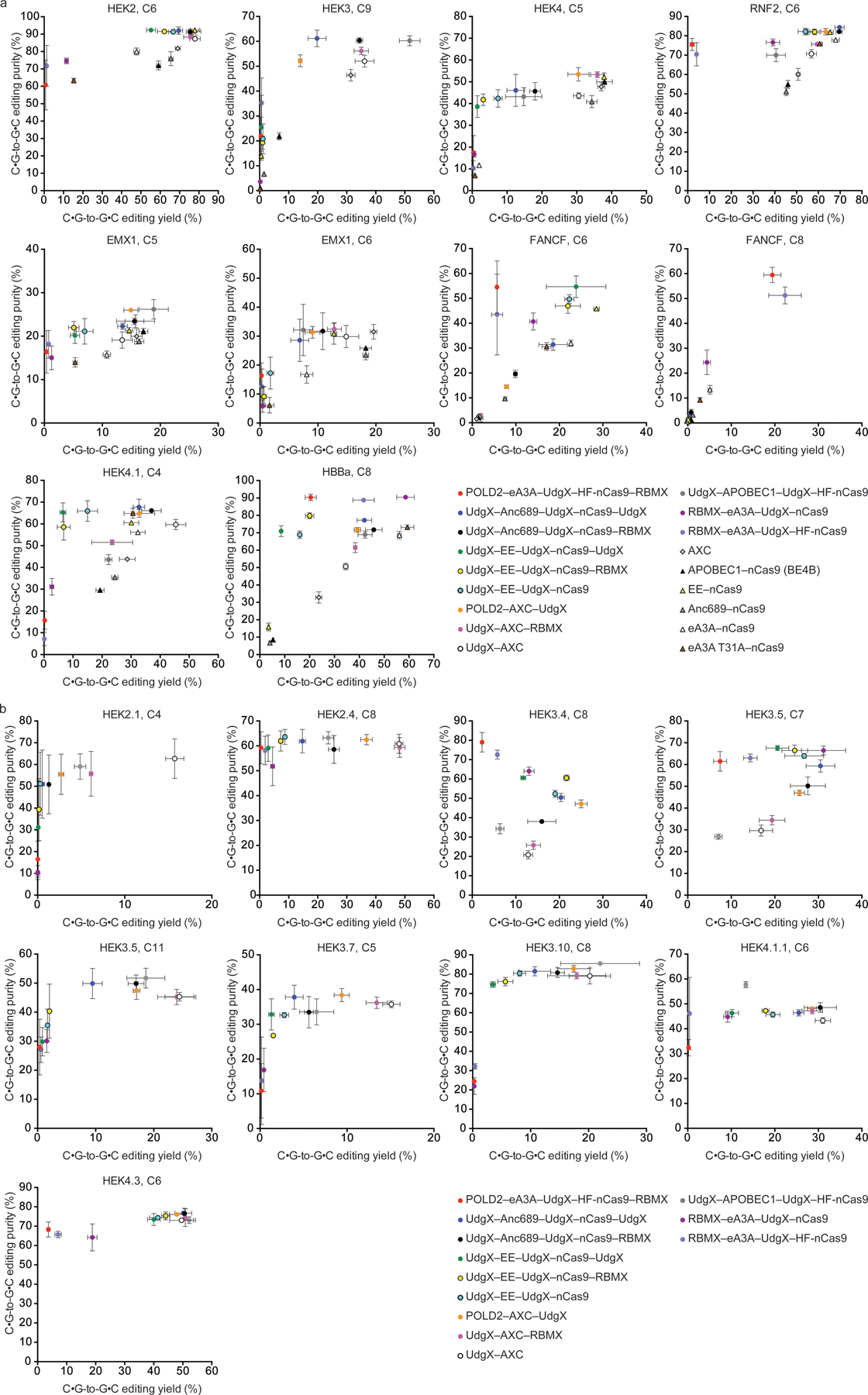
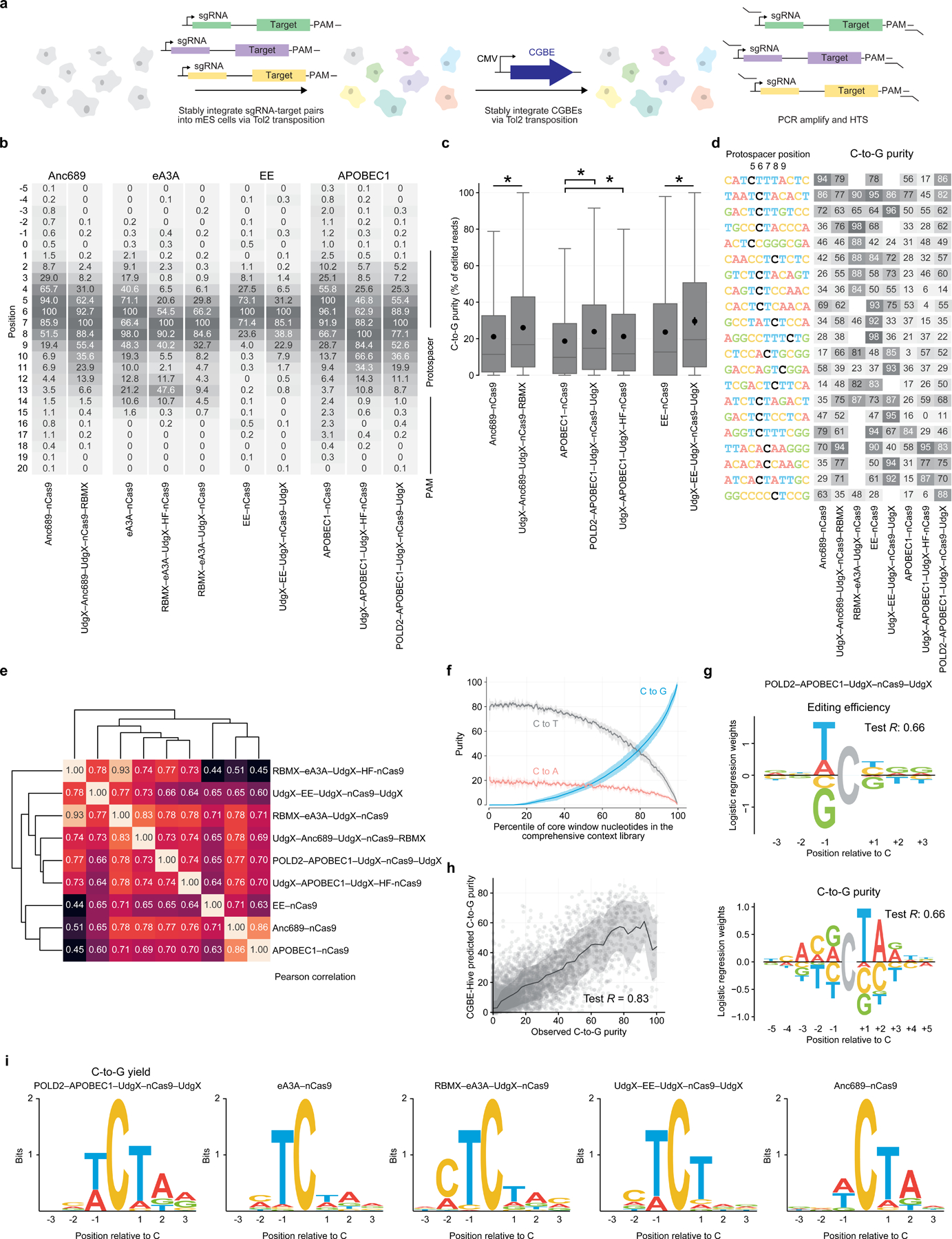
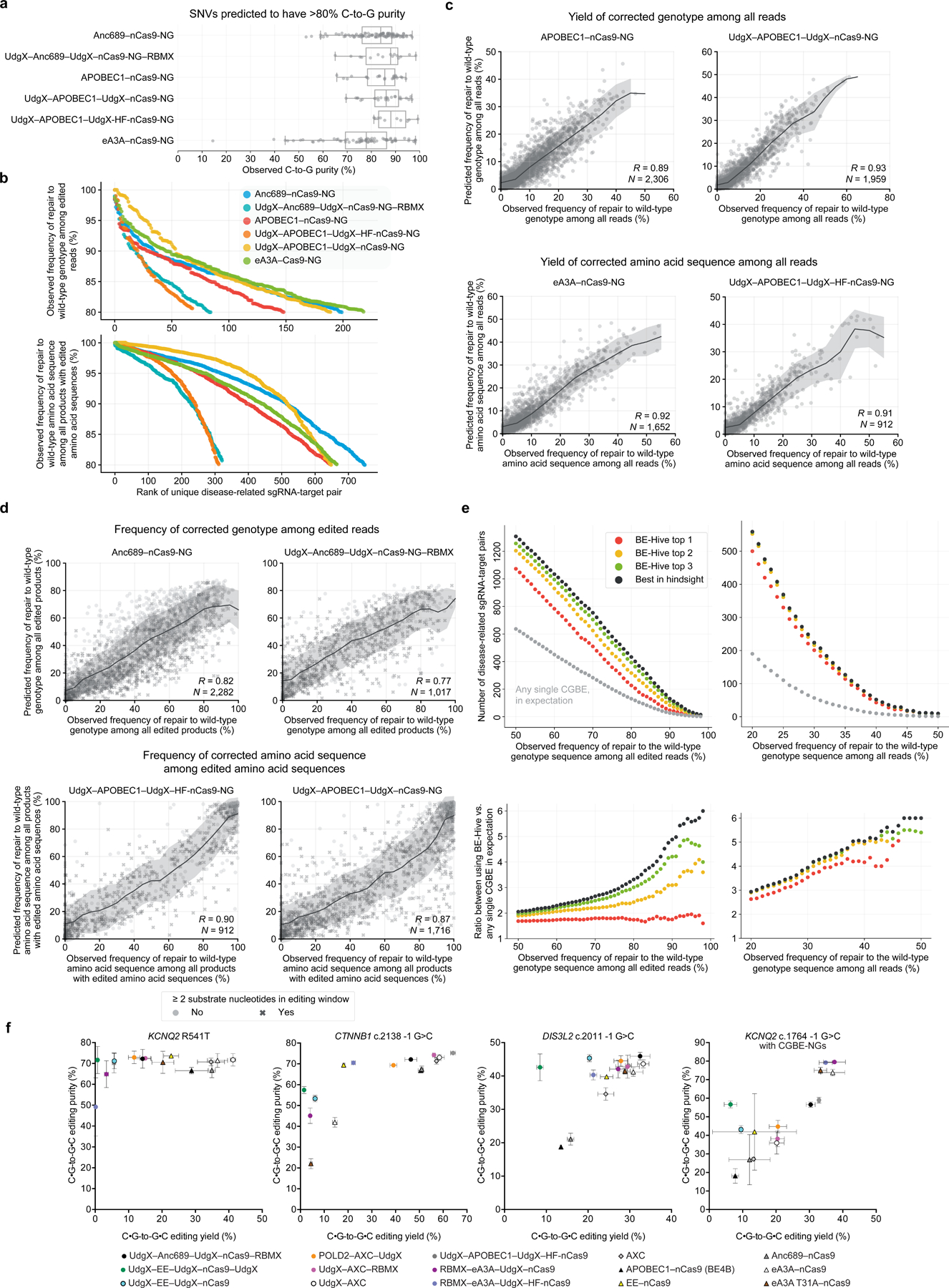
Comment in
-
Improving CRISPR tools by elucidating DNA repair.Nat Biotechnol. 2021 Dec;39(12):1512-1514. doi: 10.1038/s41587-021-01149-2. Nat Biotechnol. 2021. PMID: 34873327 No abstract available.
References
-
- Nishida K et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science 353, aaf8729–aaf8729 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
