

Causal role for the primate superior colliculus in the computation of evidence for perceptual decisions
- PMID: 34183869
- PMCID: PMC8338902
- DOI: 10.1038/s41593-021-00878-6
Causal role for the primate superior colliculus in the computation of evidence for perceptual decisions
Abstract
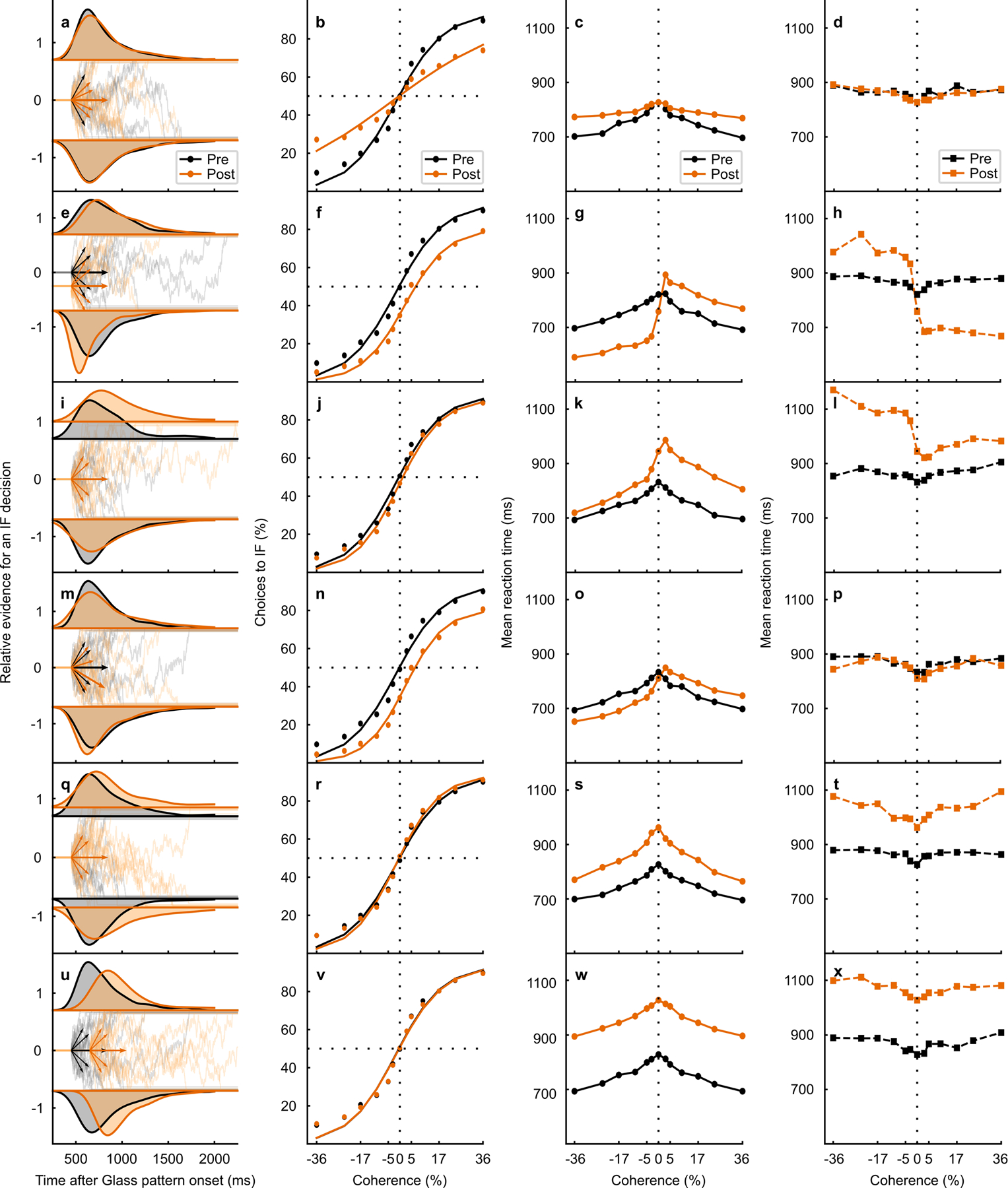
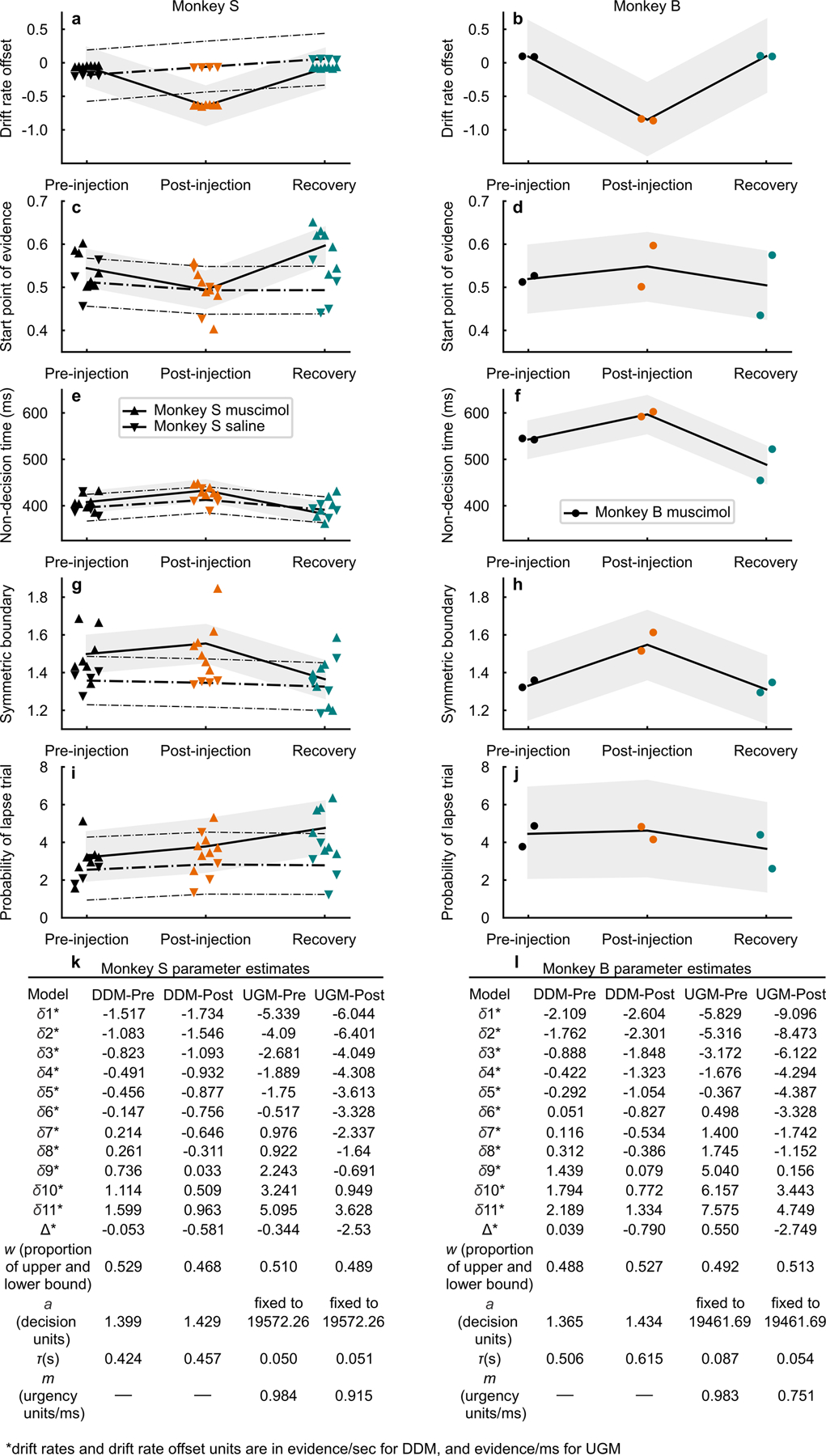
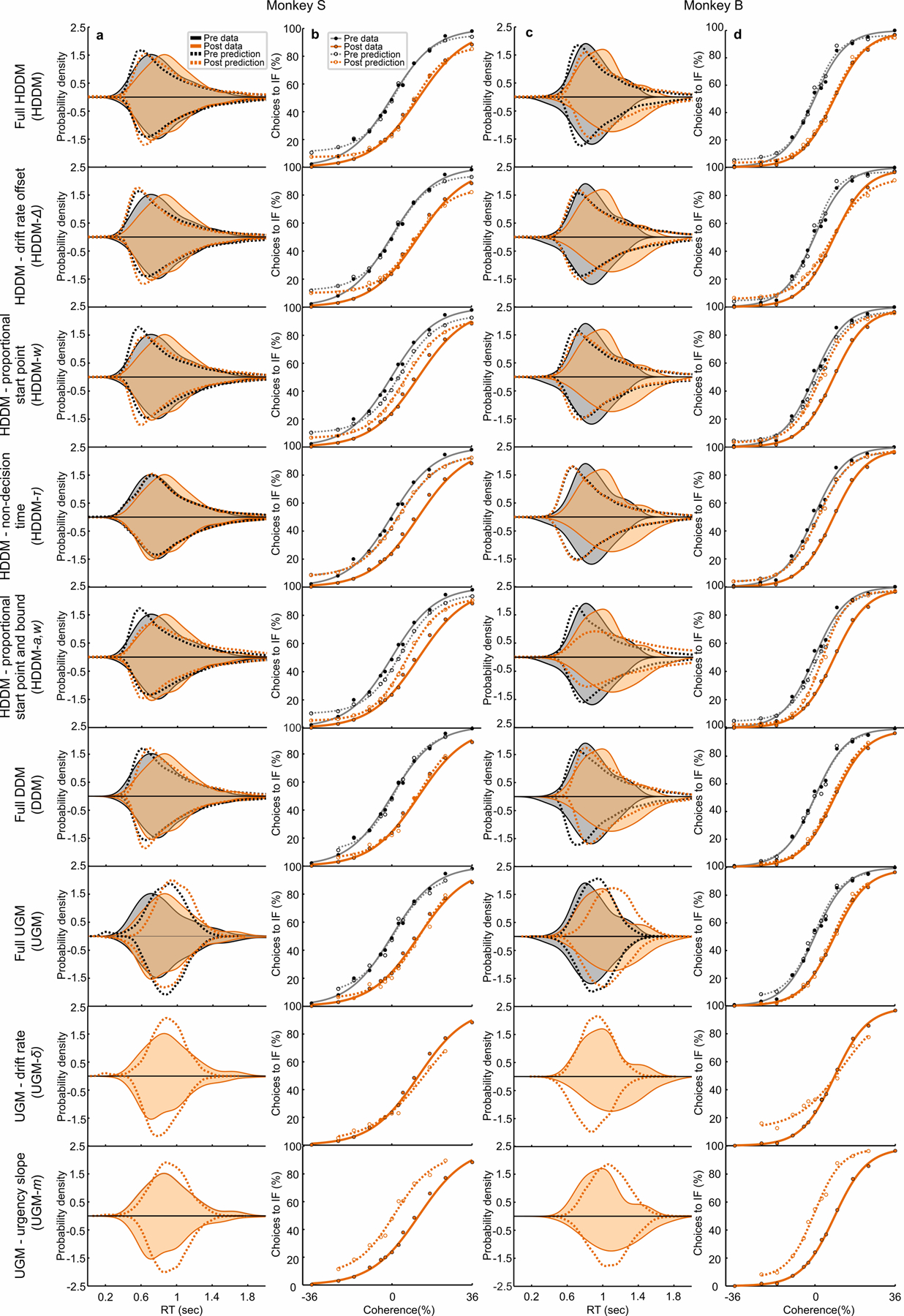
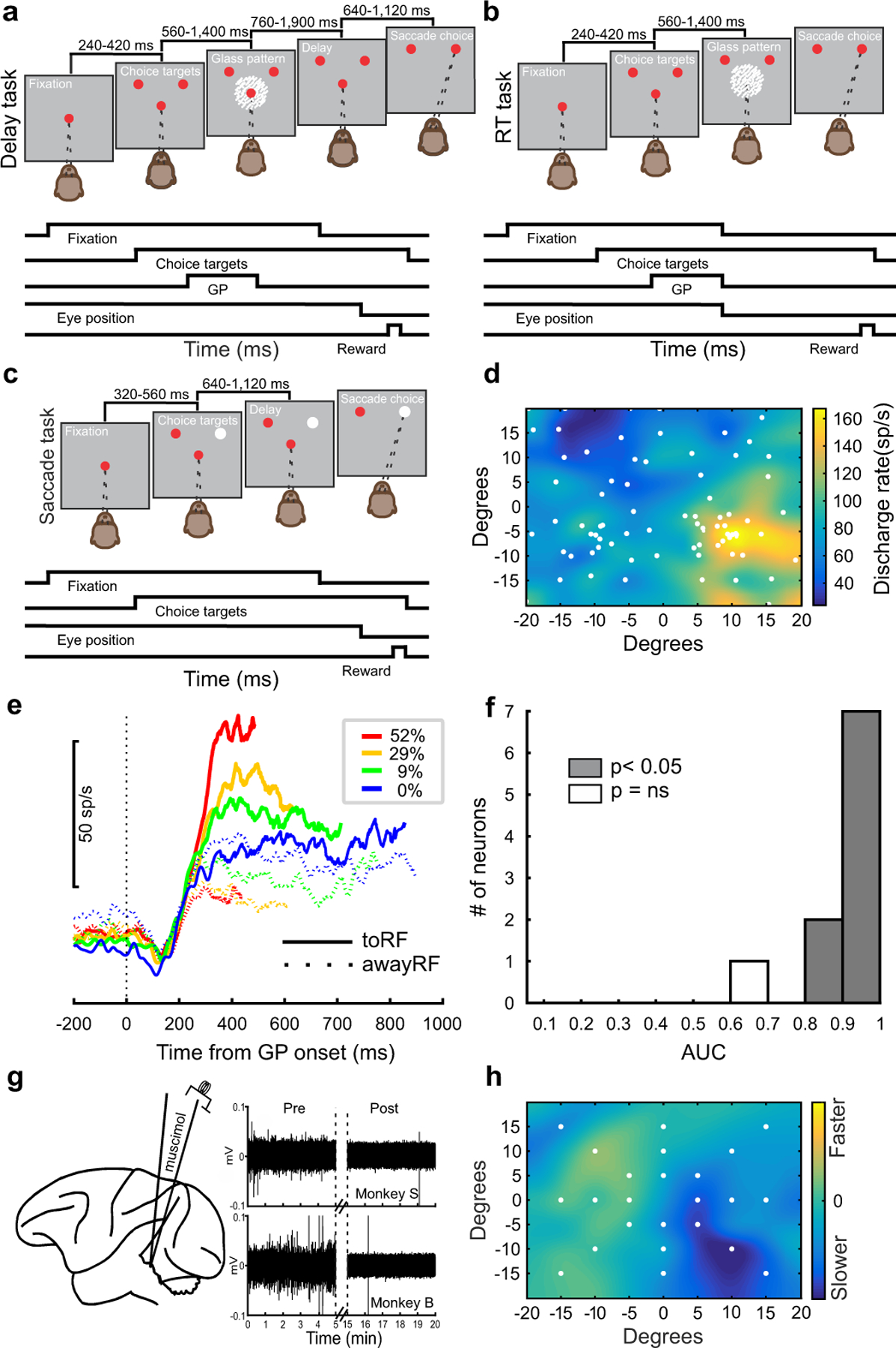
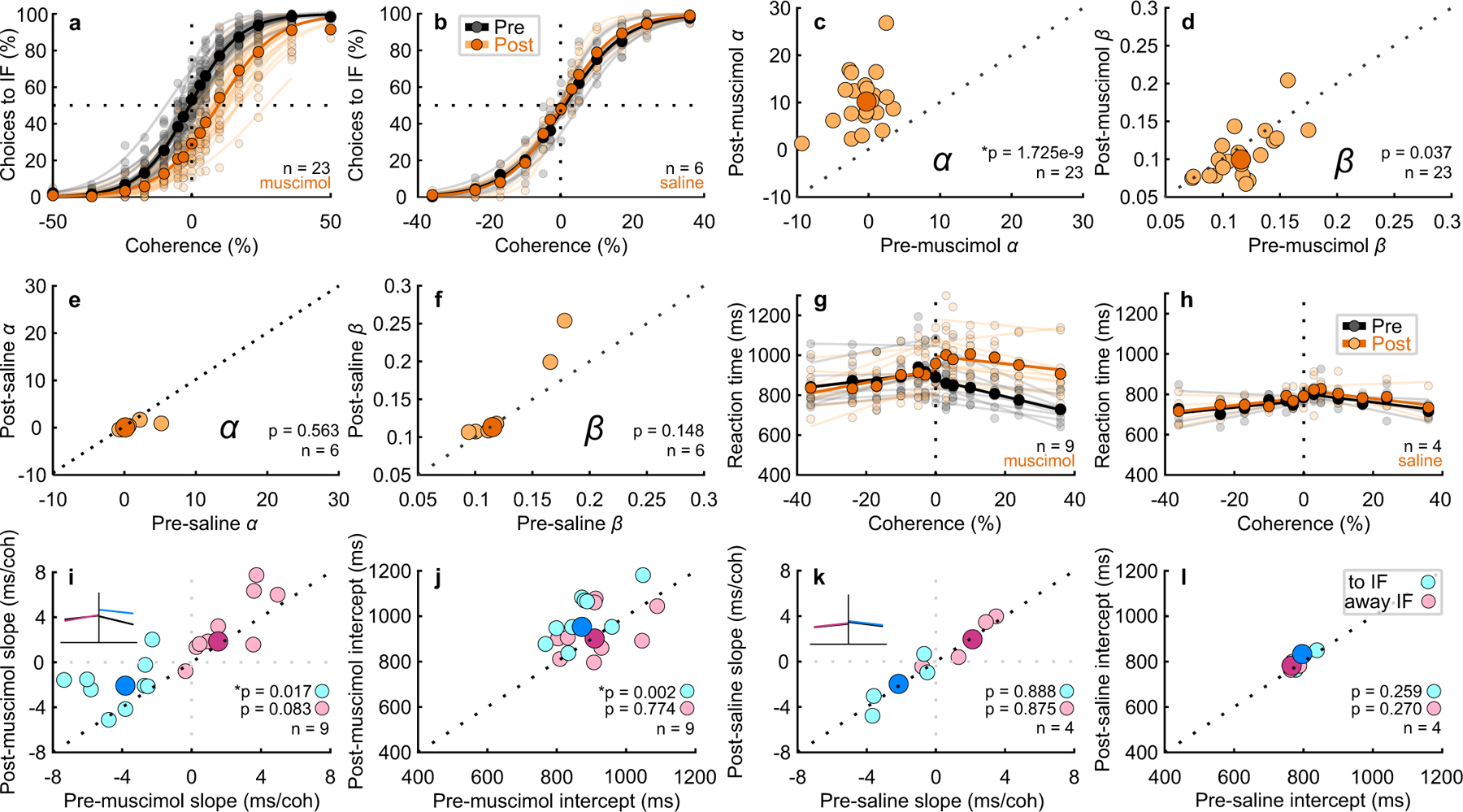
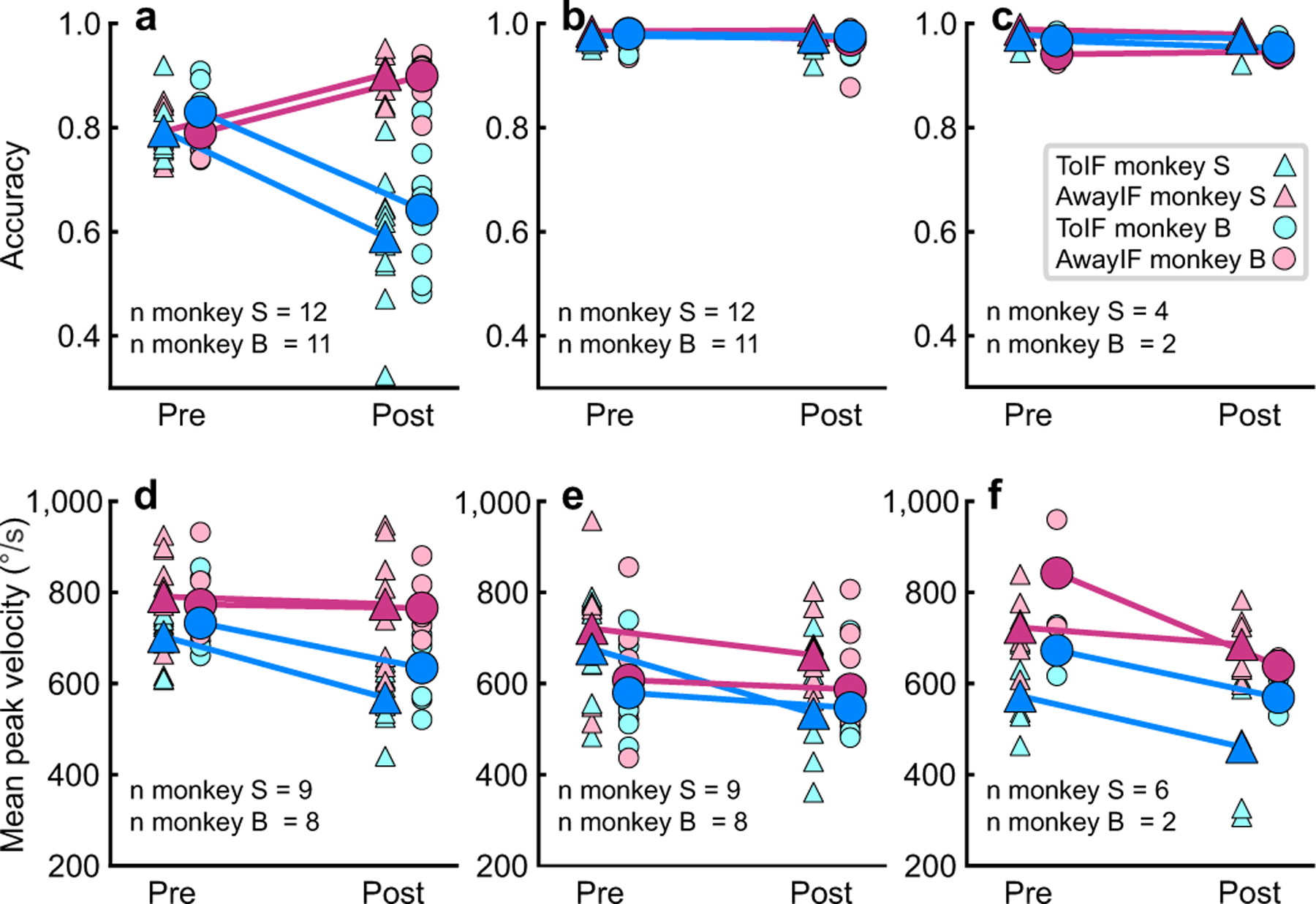
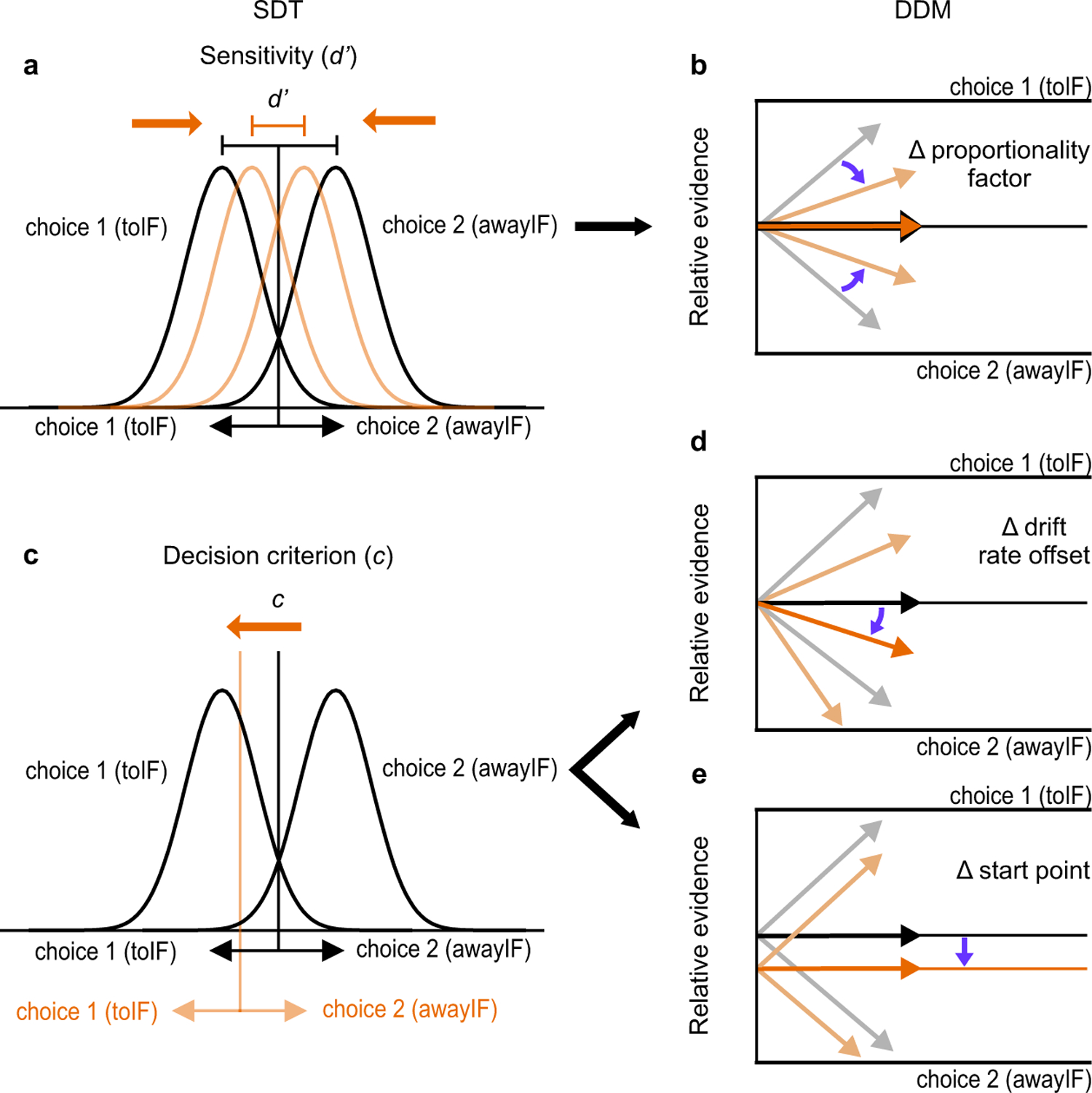
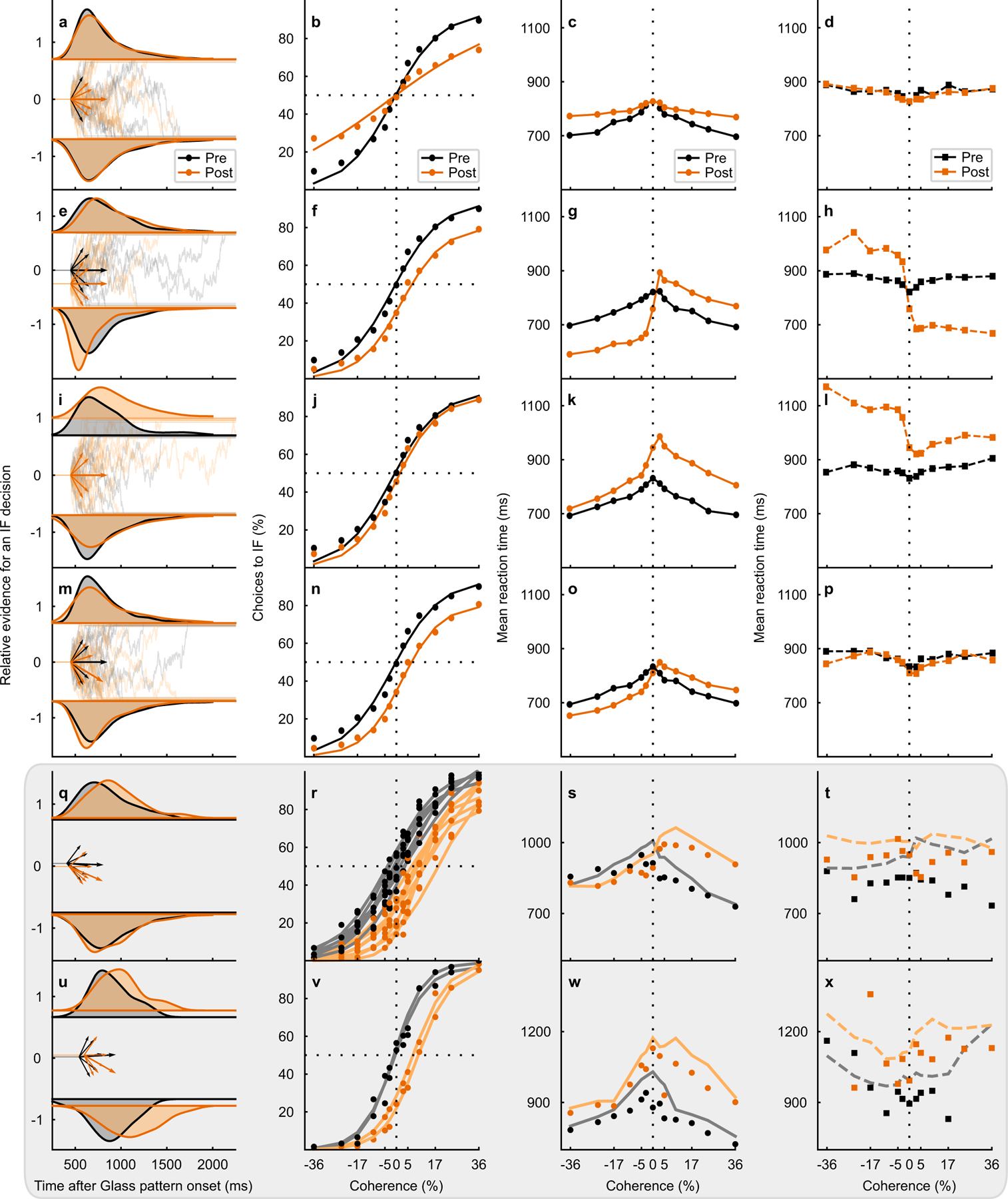
Trained monkeys performed a two-choice perceptual decision-making task in which they reported the perceived orientation of a dynamic Glass pattern, before and after unilateral, reversible, inactivation of a brainstem area-the superior colliculus (SC)-involved in preparing eye movements. We found that unilateral SC inactivation produced significant decision biases and changes in reaction times consistent with a causal role for the primate SC in perceptual decision-making. Fitting signal detection theory and sequential sampling models to the data showed that SC inactivation produced a decrease in the relative evidence for contralateral decisions, as if adding a constant offset to a time-varying evidence signal for the ipsilateral choice. The results provide causal evidence for an embodied cognition model of perceptual decision-making and provide compelling evidence that the SC of primates (a brainstem structure) plays a causal role in how evidence is computed for decisions-a process usually attributed to the forebrain.
© 2021. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Figures
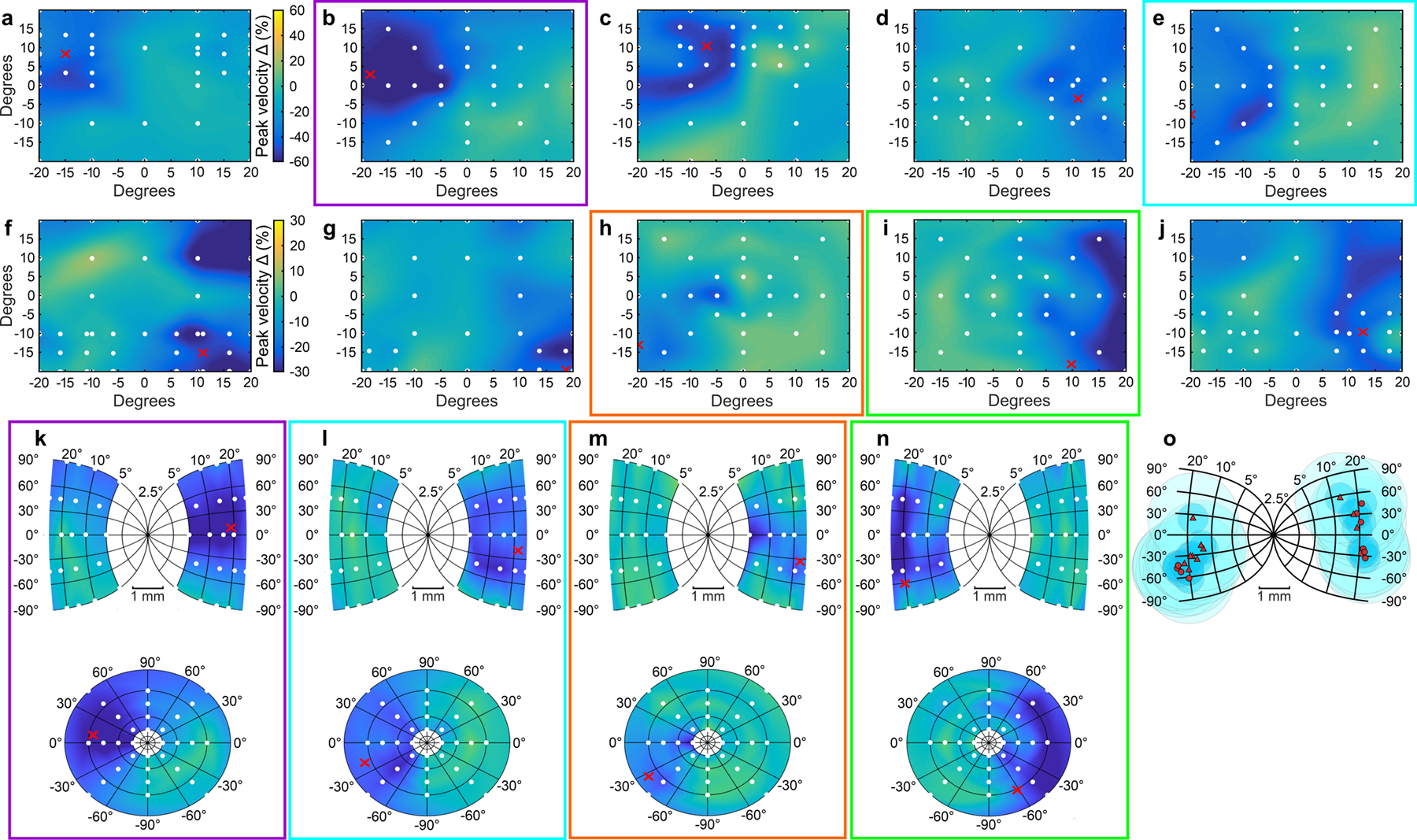
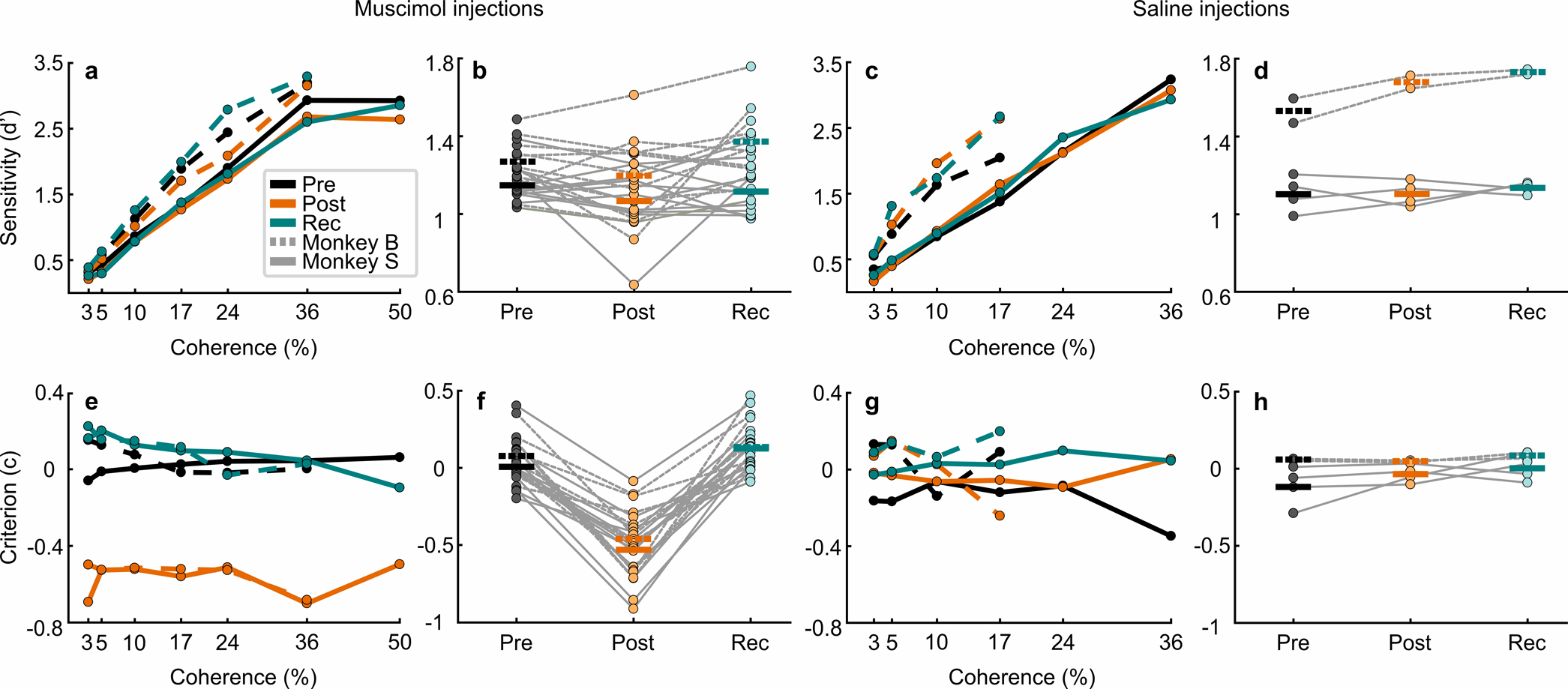
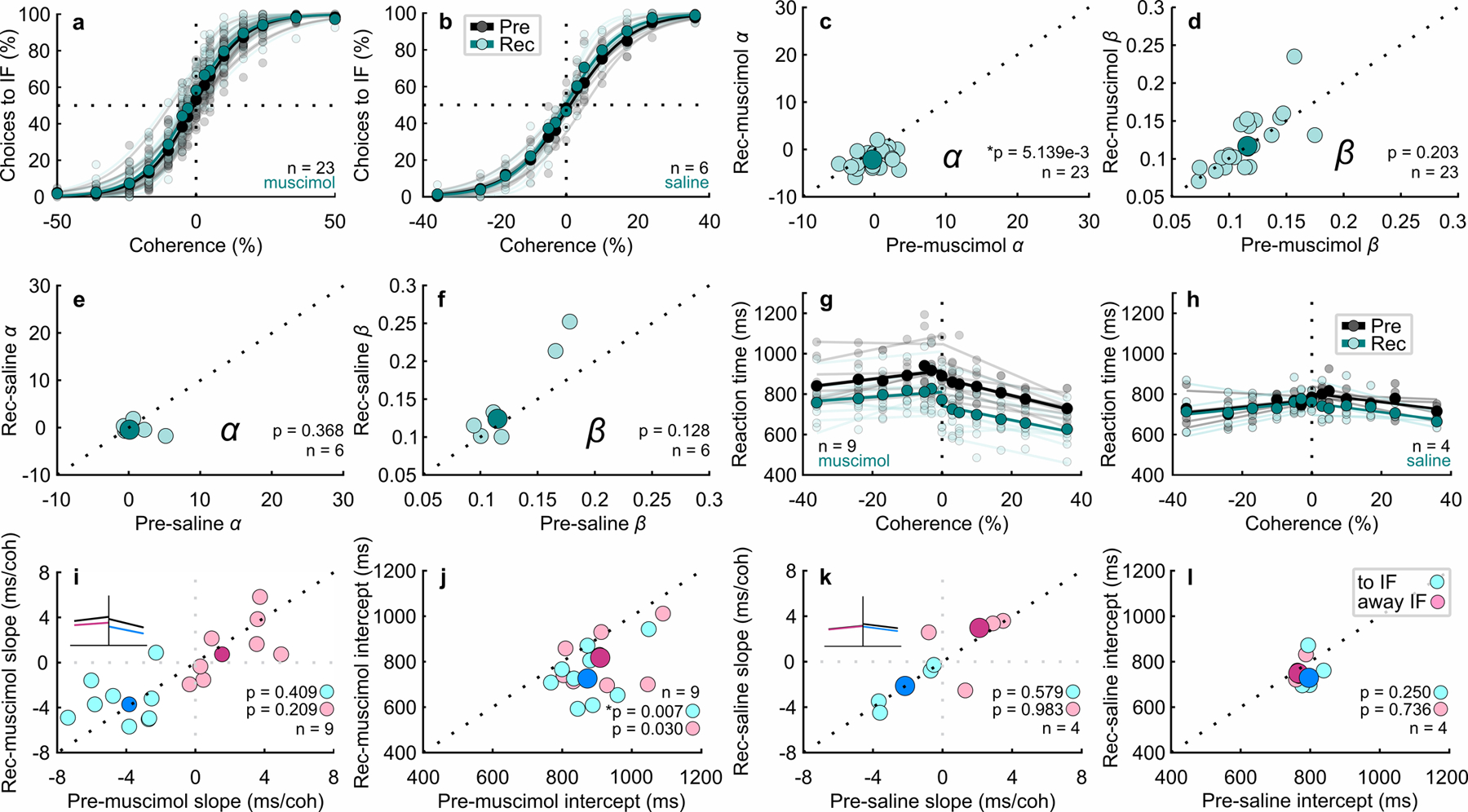
Comment in
-
Superior colliculus activates new perspectives on decision-making.Nat Neurosci. 2021 Aug;24(8):1048-1050. doi: 10.1038/s41593-021-00885-7. Nat Neurosci. 2021. PMID: 34183868 No abstract available.
References
-
- Kim JN & Shadlen MN Neural correlates of a decision in the dorsolateral prefrontal cortex of the macaque. Nature Neuroscience. 2, 176–185 (1999). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
