

Self-Organization of Cellular Units
- PMID: 34186005
- PMCID: PMC9059766
- DOI: 10.1146/annurev-cellbio-120319-025356
Self-Organization of Cellular Units
Abstract
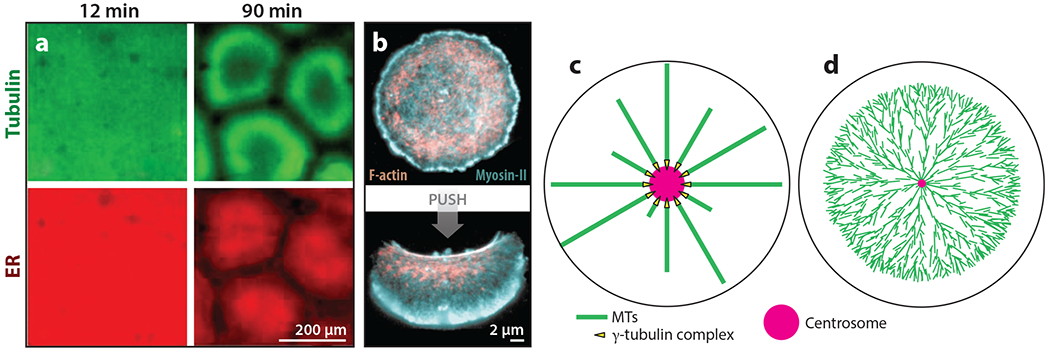
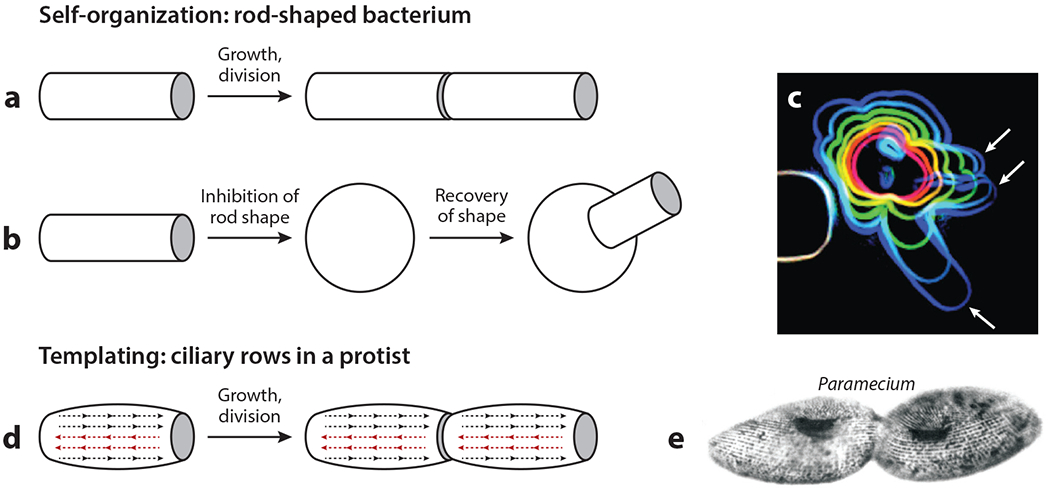
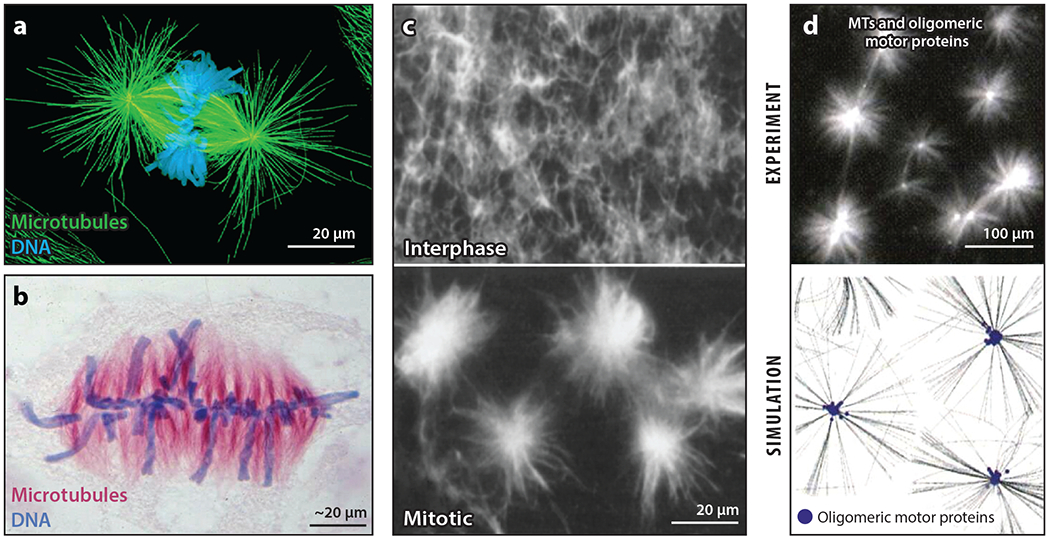
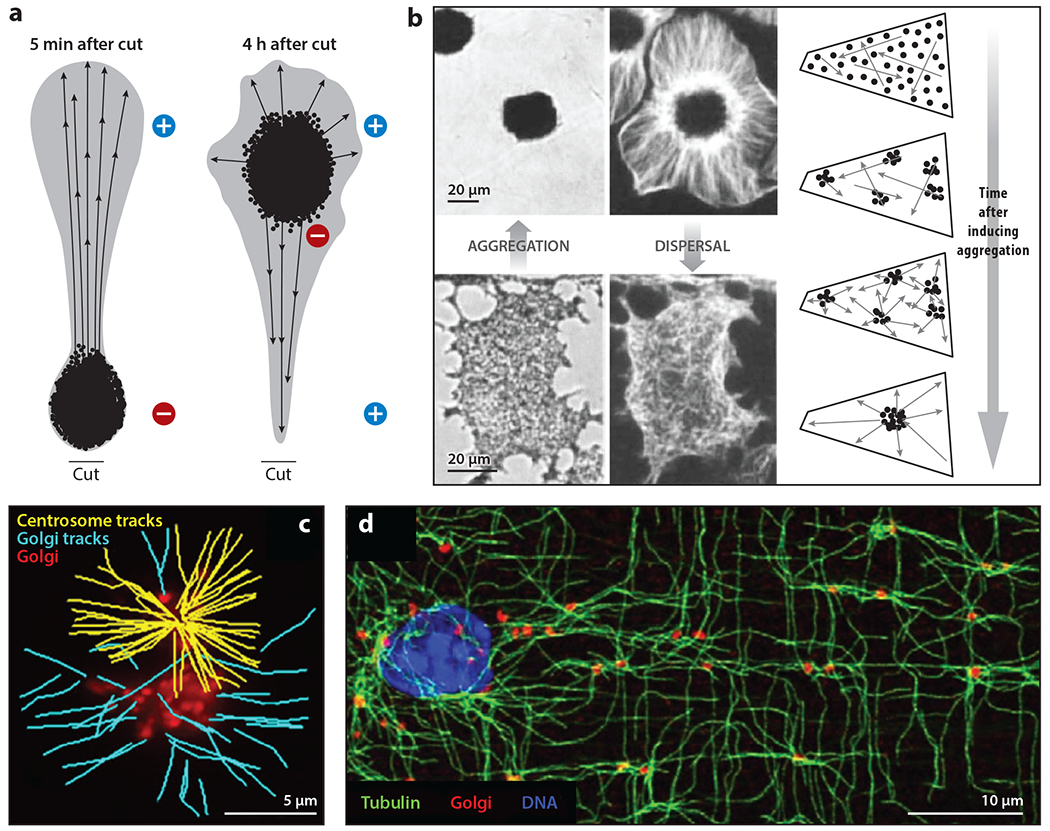
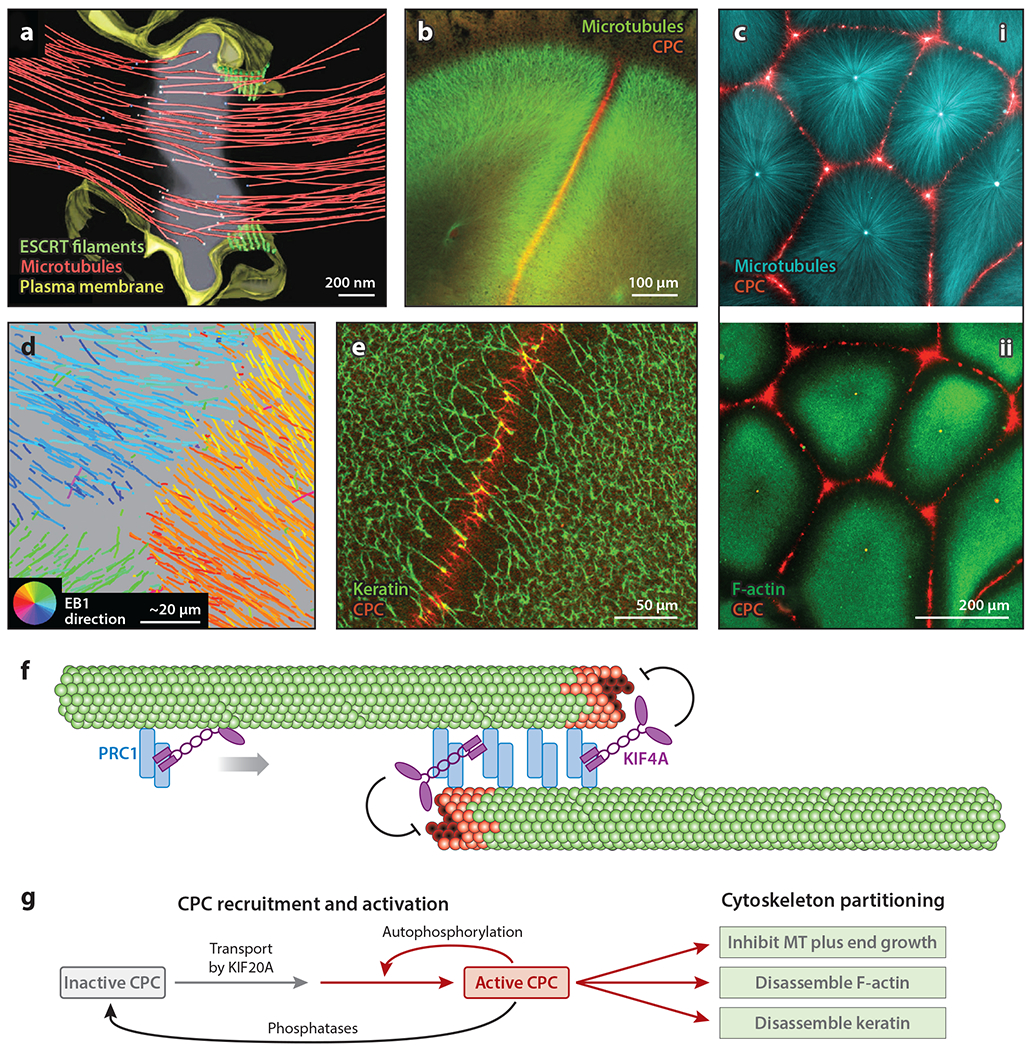
The purpose of this review is to explore self-organizing mechanisms that pattern microtubules (MTs) and spatially organize animal cell cytoplasm, inspired by recent experiments in frog egg extract. We start by reviewing conceptual distinctions between self-organizing and templating mechanisms for subcellular organization. We then discuss self-organizing mechanisms that generate radial MT arrays and cell centers in the absence of centrosomes. These include autocatalytic MT nucleation, transport of minus ends, and nucleation from organelles such as melanosomes and Golgi vesicles that are also dynein cargoes. We then discuss mechanisms that partition the cytoplasm in syncytia, in which multiple nuclei share a common cytoplasm, starting with cytokinesis, when all metazoan cells are transiently syncytial. The cytoplasm of frog eggs is partitioned prior to cytokinesis by two self-organizing modules, protein regulator of cytokinesis 1 (PRC1)-kinesin family member 4A (KIF4A) and chromosome passenger complex (CPC)-KIF20A. Similar modules may partition longer-lasting syncytia, such as early Drosophila embryos. We end by discussing shared mechanisms and principles for the MT-based self-organization of cellular units.
Keywords: centrosome; cytoskeleton; microtubules; nucleation; self-organization; syncytium.
Figures
References
-
- Adriaans IE, Hooikaas PJ, Aher A, Vromans MJM, van Es RM, et al. 2020. MKLP2 is a motile kinesin that transports the chromosomal passenger complex during anaphase. Curr. Biol 30:2628–37.e9 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
