

Toxoplasma gondii Toxolysin 4 Contributes to Efficient Parasite Egress from Host Cells
- PMID: 34190588
- PMCID: PMC8265663
- DOI: 10.1128/mSphere.00444-21
Toxoplasma gondii Toxolysin 4 Contributes to Efficient Parasite Egress from Host Cells
Abstract
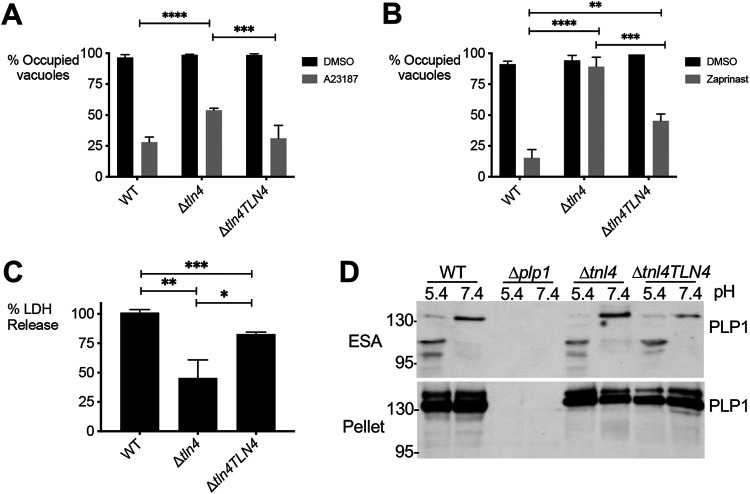
Egress from host cells is an essential step in the lytic cycle of T. gondii and other apicomplexan parasites; however, only a few parasite secretory proteins are known to affect this process. The putative metalloproteinase toxolysin 4 (TLN4) was previously shown to be an extensively processed microneme protein, but further characterization was impeded by the inability to genetically ablate TLN4. Here, we show that TLN4 has the structural properties of an M16 family metalloproteinase, that it possesses proteolytic activity on a model substrate, and that genetic disruption of TLN4 reduces the efficiency of egress from host cells. Complementation of the knockout strain with the TLN4 coding sequence significantly restored egress competency, affirming that the phenotype of the Δtln4 parasite was due to the absence of TLN4. This work identifies TLN4 as the first metalloproteinase and the second microneme protein to function in T. gondii egress. The study also lays a foundation for future mechanistic studies defining the precise role of TLN4 in parasite exit from host cells. IMPORTANCE After replicating within infected host cells, the single-celled parasite Toxoplasma gondii must rupture out of such cells in a process termed egress. Although it is known that T. gondii egress is an active event that involves disruption of host-derived membranes surrounding the parasite, very few proteins that are released by the parasite are known to facilitate egress. In this study, we identify a parasite secretory protease that is necessary for efficient and timely egress, laying the foundation for understanding precisely how this protease facilitates T. gondii exit from host cells.
Keywords: Toxoplasma gondii; apicomplexan parasites; egress; intracellular parasites; proteases.
Figures
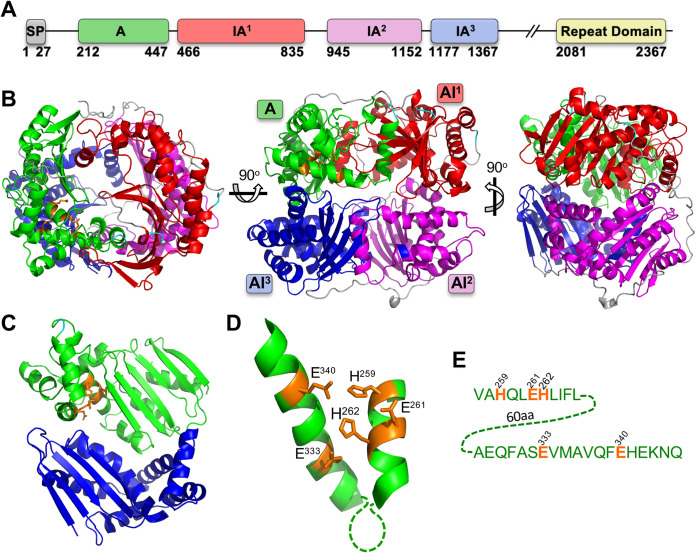
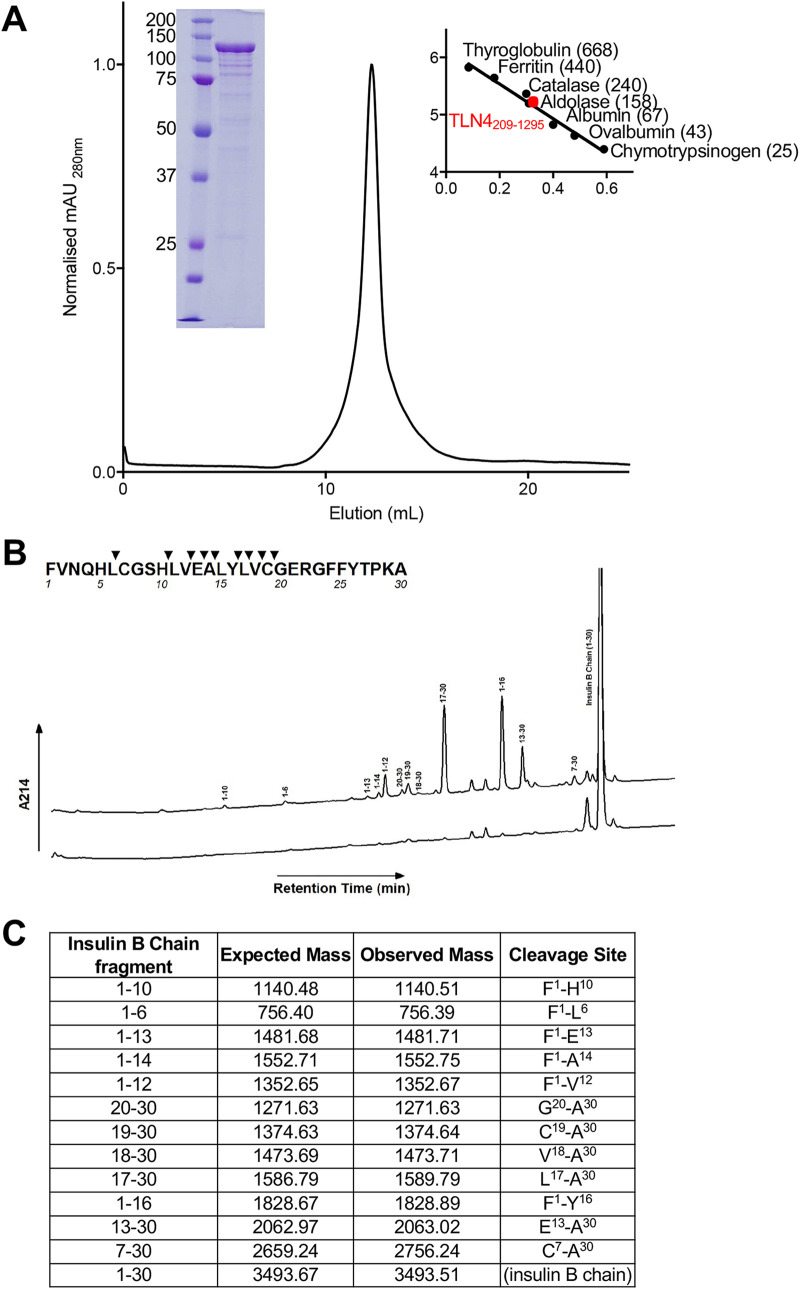
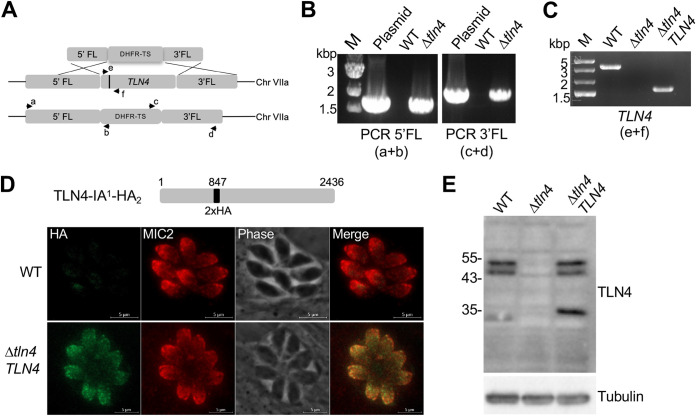
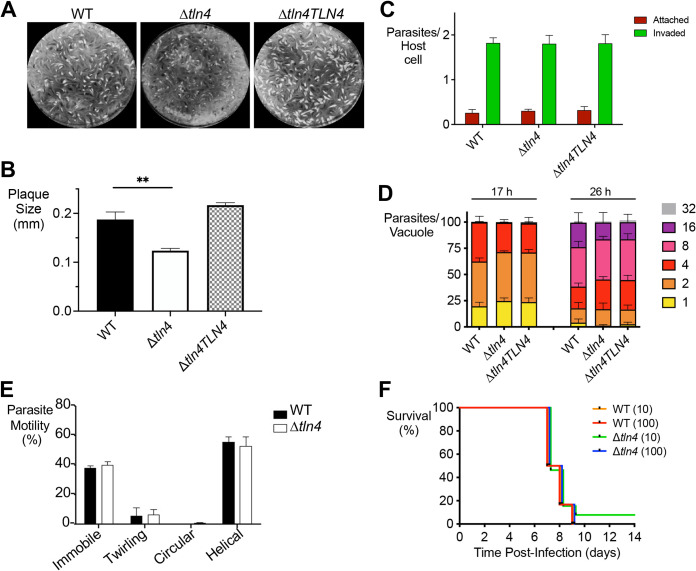
References
Grants and funding
LinkOut - more resources
Full Text Sources
