

BK channel activation by L-type Ca2+ channels CaV1.2 and CaV1.3 during the subthreshold phase of an action potential
- PMID: 34191630
- PMCID: PMC8409951
- DOI: 10.1152/jn.00089.2021
BK channel activation by L-type Ca2+ channels CaV1.2 and CaV1.3 during the subthreshold phase of an action potential
Abstract
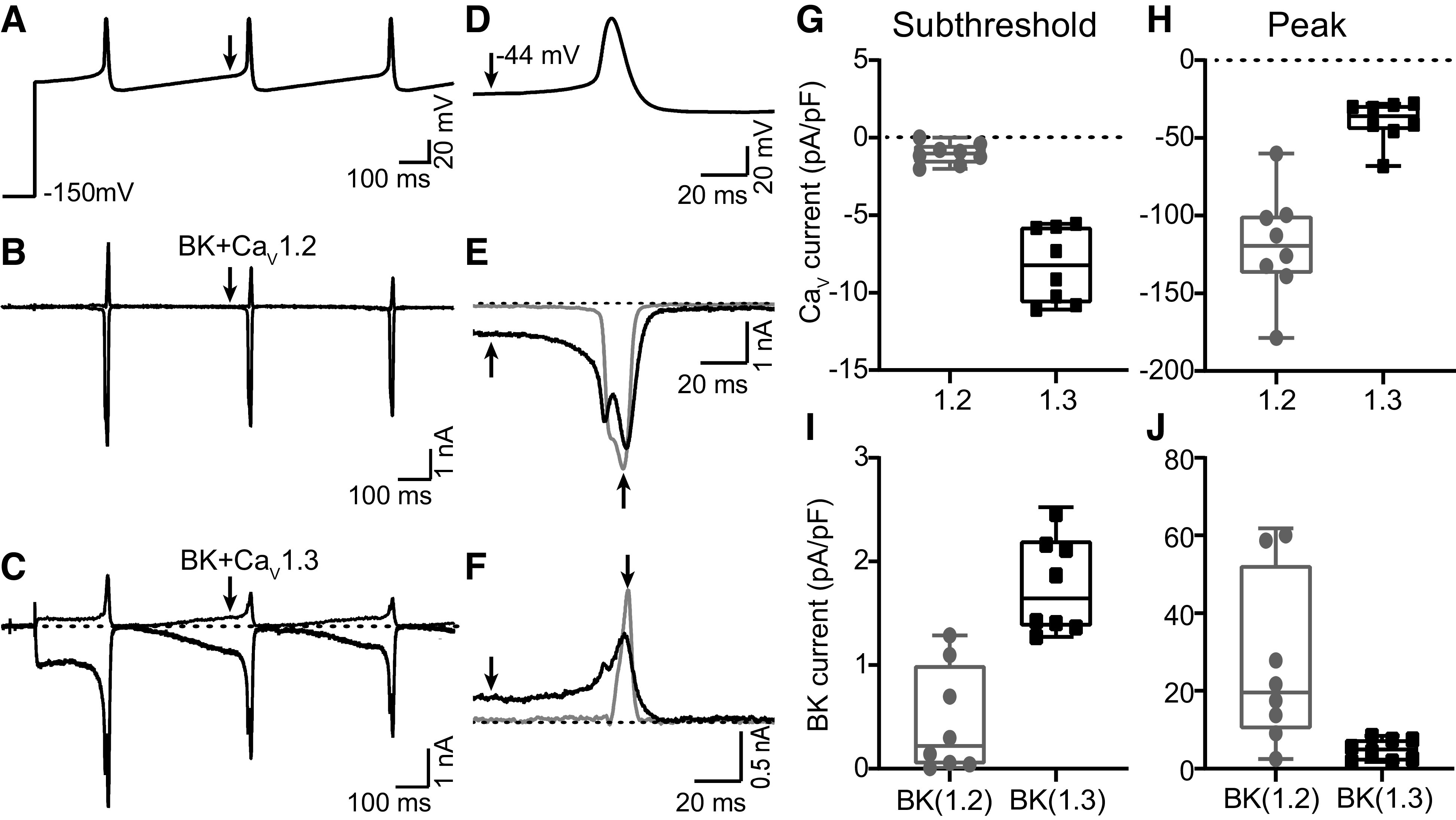
Mammalian circadian (24 h) rhythms are timed by the pattern of spontaneous action potential firing in the suprachiasmatic nucleus (SCN). This oscillation in firing is produced through circadian regulation of several membrane currents, including large-conductance Ca2+- and voltage-activated K+ (BK) and L-type Ca2+ channel (LTCC) currents. During the day steady-state BK currents depend mostly on LTCCs for activation, whereas at night they depend predominantly on ryanodine receptors (RyRs). However, the contribution of these Ca2+ channels to BK channel activation during action potential firing has not been thoroughly investigated. In this study, we used a pharmacological approach to determine that both LTCCs and RyRs contribute to the baseline membrane potential of SCN action potential waveforms, as well as action potential-evoked BK current, during the day and night, respectively. Since the baseline membrane potential is a major determinant of circadian firing rate, we focused on the LTCCs contributing to low voltage activation of BK channels during the subthreshold phase. For these experiments, two LTCC subtypes found in SCN (CaV1.2 and CaV1.3) were coexpressed with BK channels in heterologous cells, where their differential contributions could be separately measured. CaV1.3 channels produced currents that were shifted to more hyperpolarized potentials compared with CaV1.2, resulting in increased subthreshold Ca2+ and BK currents during an action potential command. These results show that although multiple Ca2+ sources in SCN can contribute to the activation of BK current during an action potential, specific BK-CaV1.3 partnerships may optimize the subthreshold BK current activation that is critical for firing rate regulation.NEW & NOTEWORTHY BK K+ channels are important regulators of firing. Although Ca2+ channels are required for their activation in excitable cells, it is not well understood how BK channels activate using these Ca2+ sources during an action potential. This study demonstrates the differences in BK current activated by CaV1.2 and CaV1.3 channels in clock neurons and heterologous cells. The results define how specific ion channel partnerships can be engaged during distinct phases of the action potential.
Keywords: BK channel; KCNMA1; L-type calcium channel; action potential; suprachiasmatic nucleus.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures




Similar articles
-
Contributions of CaV1.3 Channels to Ca2+ Current and Ca2+-Activated BK Current in the Suprachiasmatic Nucleus.Front Physiol. 2021 Sep 28;12:737291. doi: 10.3389/fphys.2021.737291. eCollection 2021. Front Physiol. 2021. PMID: 34650447 Free PMC article.
-
Loss of Cav1.3 channels reveals the critical role of L-type and BK channel coupling in pacemaking mouse adrenal chromaffin cells.J Neurosci. 2010 Jan 13;30(2):491-504. doi: 10.1523/JNEUROSCI.4961-09.2010. J Neurosci. 2010. PMID: 20071512 Free PMC article.
-
Differential contribution of Ca2+ sources to day and night BK current activation in the circadian clock.J Gen Physiol. 2018 Feb 5;150(2):259-275. doi: 10.1085/jgp.201711945. Epub 2017 Dec 13. J Gen Physiol. 2018. PMID: 29237755 Free PMC article.
-
Ca(v)1.3 and BK channels for timing and regulating cell firing.Mol Neurobiol. 2010 Dec;42(3):185-98. doi: 10.1007/s12035-010-8151-3. Epub 2010 Nov 20. Mol Neurobiol. 2010. PMID: 21088933 Review.
-
CaV1.3 as pacemaker channels in adrenal chromaffin cells: specific role on exo- and endocytosis?Channels (Austin). 2010 Nov-Dec;4(6):440-6. doi: 10.4161/chan.4.6.12866. Epub 2010 Nov 1. Channels (Austin). 2010. PMID: 21084859 Review.
Cited by
-
Evidence for Dual Activation of IK(M) and IK(Ca) Caused by QO-58 (5-(2,6-Dichloro-5-fluoropyridin-3-yl)-3-phenyl-2-(trifluoromethyl)-1H-pyrazolol[1,5-a]pyrimidin-7-one).Int J Mol Sci. 2022 Jun 24;23(13):7042. doi: 10.3390/ijms23137042. Int J Mol Sci. 2022. PMID: 35806047 Free PMC article.
-
T-Type Voltage-Gated Calcium Channels: Potential Regulators of Smooth Muscle Contractility.Int J Mol Sci. 2024 Nov 19;25(22):12420. doi: 10.3390/ijms252212420. Int J Mol Sci. 2024. PMID: 39596484 Free PMC article. Review.
-
Mechanisms and physiological implications of cooperative gating of clustered ion channels.Physiol Rev. 2022 Jul 1;102(3):1159-1210. doi: 10.1152/physrev.00022.2021. Epub 2021 Dec 20. Physiol Rev. 2022. PMID: 34927454 Free PMC article. Review.
-
Kinesin-1 mediates proper ER folding of the CaV1.2 channel and maintains mouse glucose homeostasis.EMBO Rep. 2024 Nov;25(11):4777-4802. doi: 10.1038/s44319-024-00246-y. Epub 2024 Sep 25. EMBO Rep. 2024. PMID: 39322740 Free PMC article.
-
Activity dependent Clustering of Neuronal L-Type Calcium Channels by CaMKII.bioRxiv [Preprint]. 2025 Jan 8:2025.01.08.631979. doi: 10.1101/2025.01.08.631979. bioRxiv. 2025. PMID: 39829809 Free PMC article. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous