

The Polycomb group protein MEDEA controls cell proliferation and embryonic patterning in Arabidopsis
- PMID: 34192526
- PMCID: PMC8279741
- DOI: 10.1016/j.devcel.2021.06.004
The Polycomb group protein MEDEA controls cell proliferation and embryonic patterning in Arabidopsis
Abstract
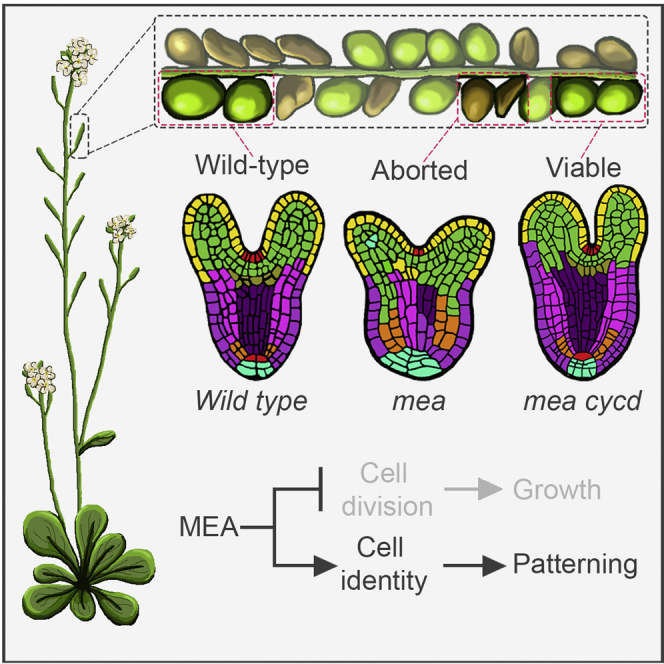
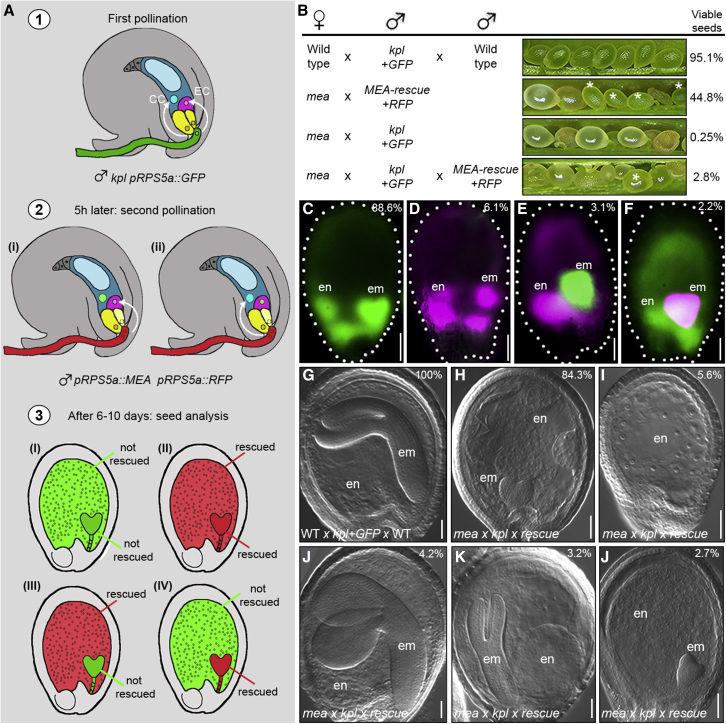
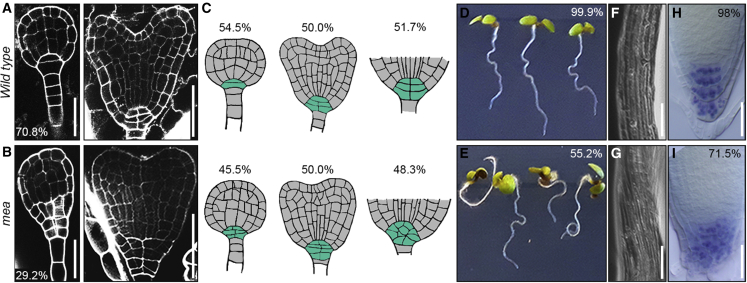
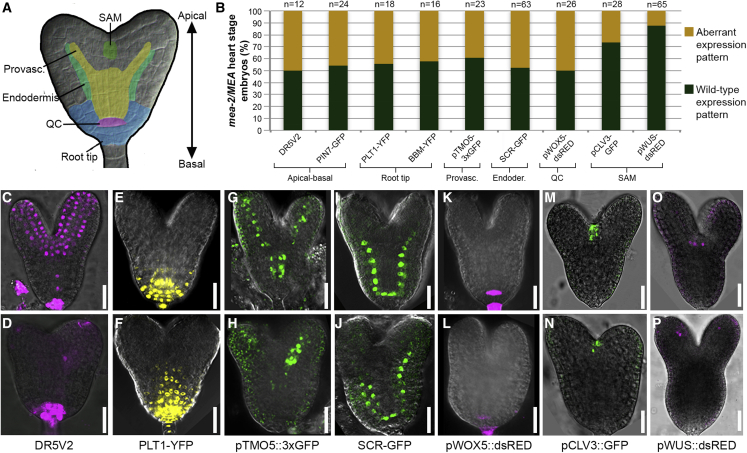
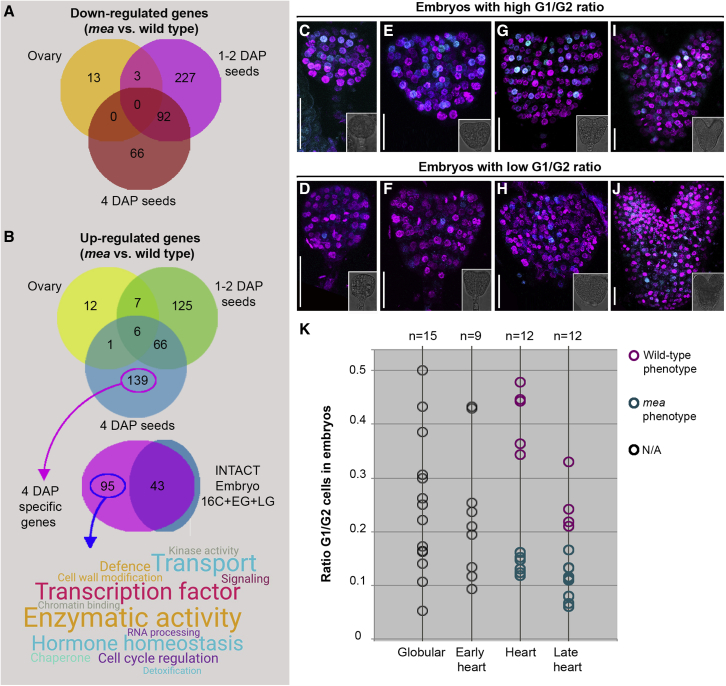
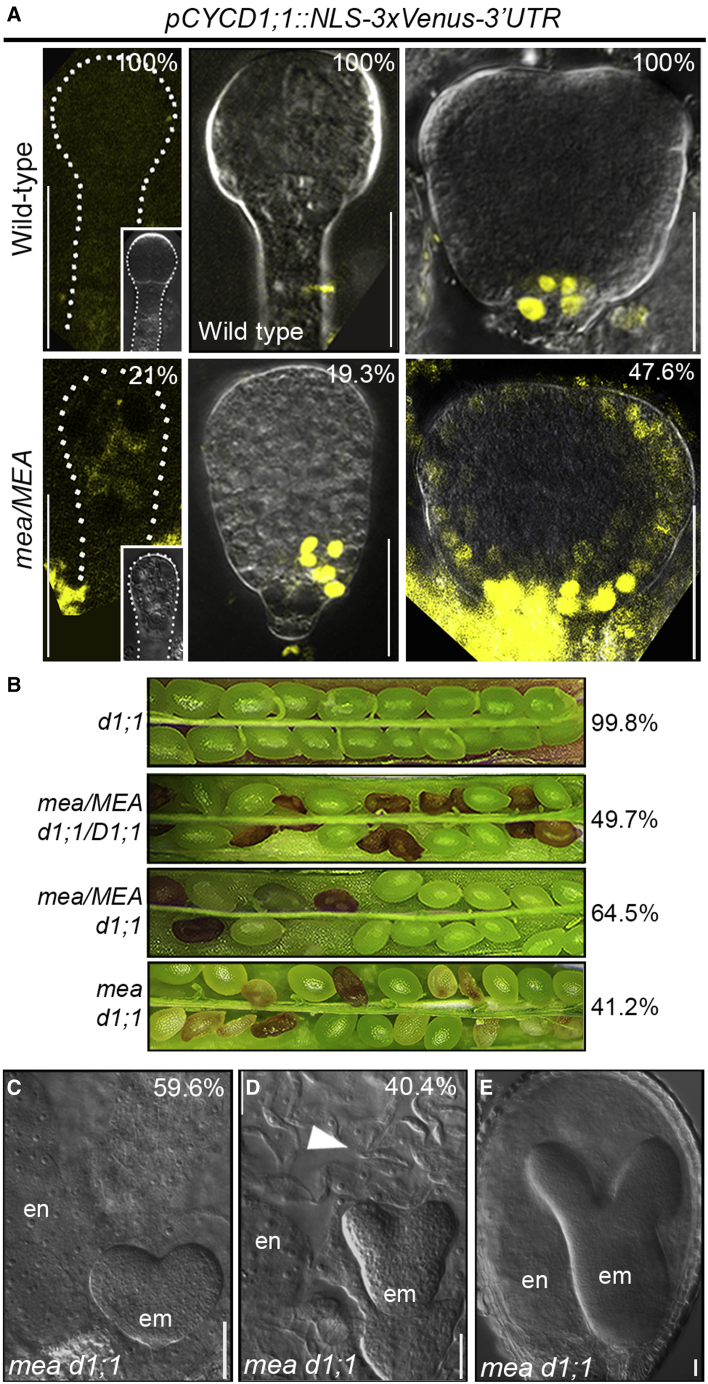
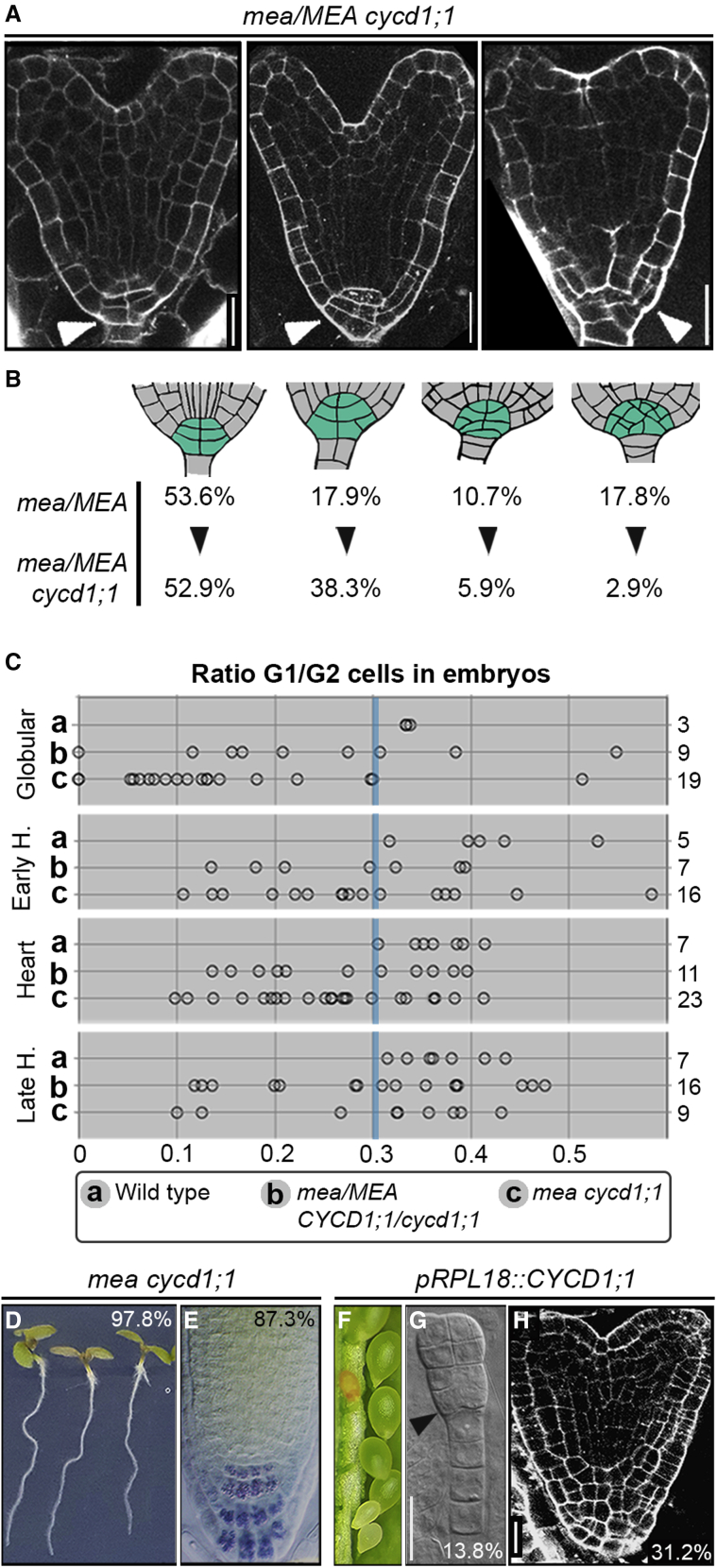
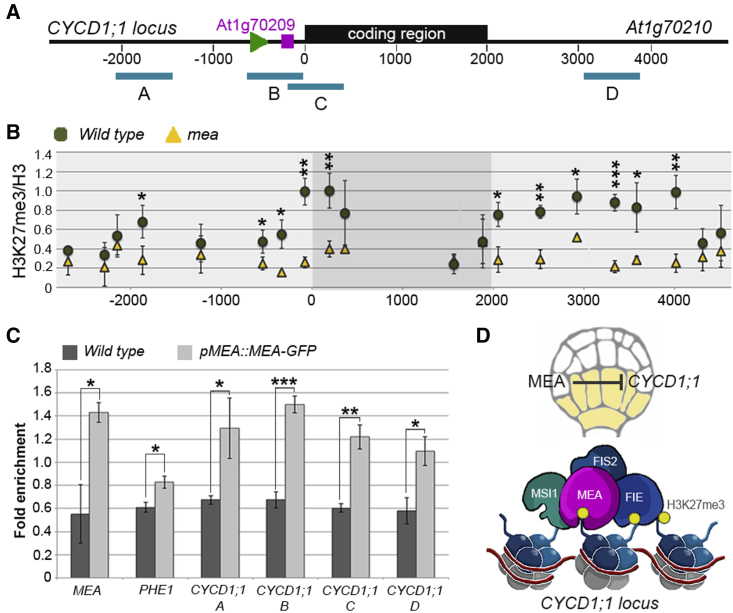
Establishing the embryonic body plan of multicellular organisms relies on precisely orchestrated cell divisions coupled with pattern formation, which, in animals, are regulated by Polycomb group (PcG) proteins. The conserved Polycomb Repressive Complex 2 (PRC2) mediates H3K27 trimethylation and comes in different flavors in Arabidopsis. The PRC2 catalytic subunit MEDEA is required for seed development; however, a role for PRC2 in embryonic patterning has been dismissed. Here, we demonstrate that embryos derived from medea eggs abort because MEDEA is required for patterning and cell lineage determination in the early embryo. Similar to PcG proteins in mammals, MEDEA regulates embryonic patterning and growth by controlling cell-cycle progression through repression of CYCD1;1, which encodes a core cell-cycle component. Thus, Arabidopsis embryogenesis is epigenetically regulated by PcG proteins, revealing that the PRC2-dependent modulation of cell-cycle progression was independently recruited to control embryonic cell proliferation and patterning in animals and plants.
Keywords: Arabidopsis; H3K27me3; MEDEA; PRC2; Polycomb group; cell proliferation; cyclin; embryonic patterning; evolutionary conservation; plant development.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
