

Orai channel C-terminal peptides are key modulators of STIM-Orai coupling and calcium signal generation
- PMID: 34192542
- PMCID: PMC8462482
- DOI: 10.1016/j.celrep.2021.109322
Orai channel C-terminal peptides are key modulators of STIM-Orai coupling and calcium signal generation
Abstract
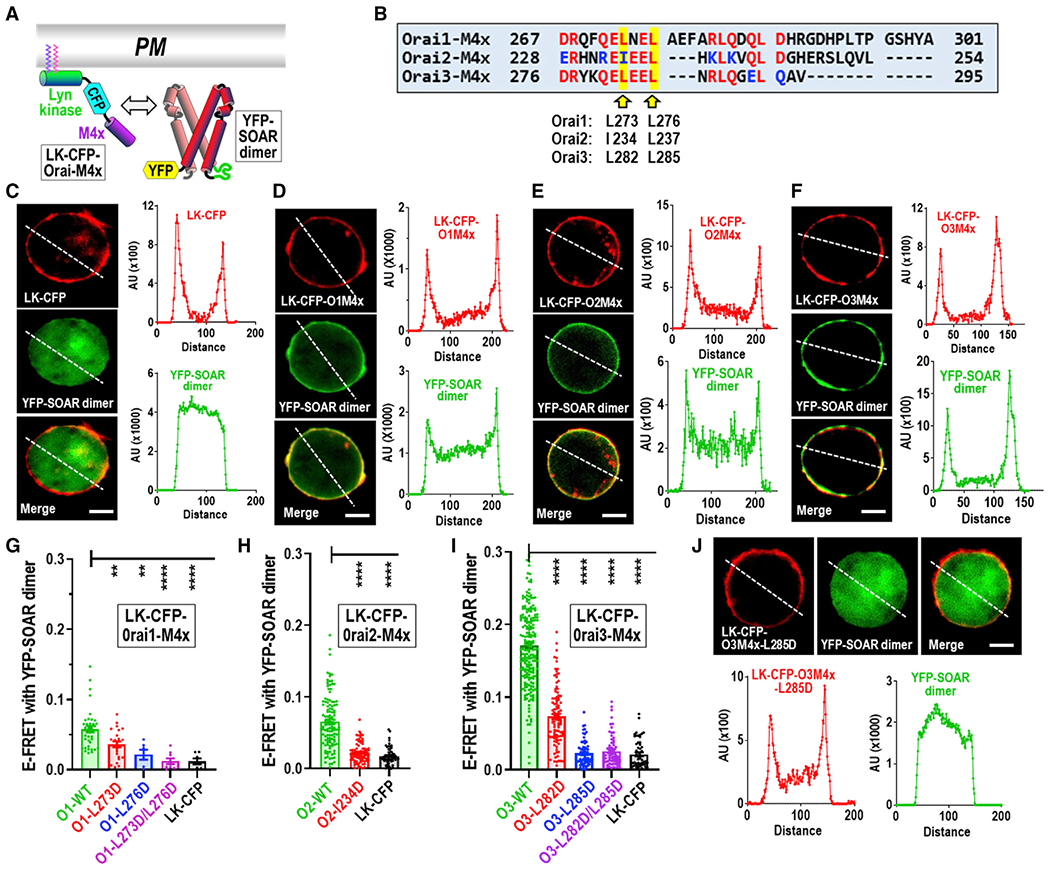
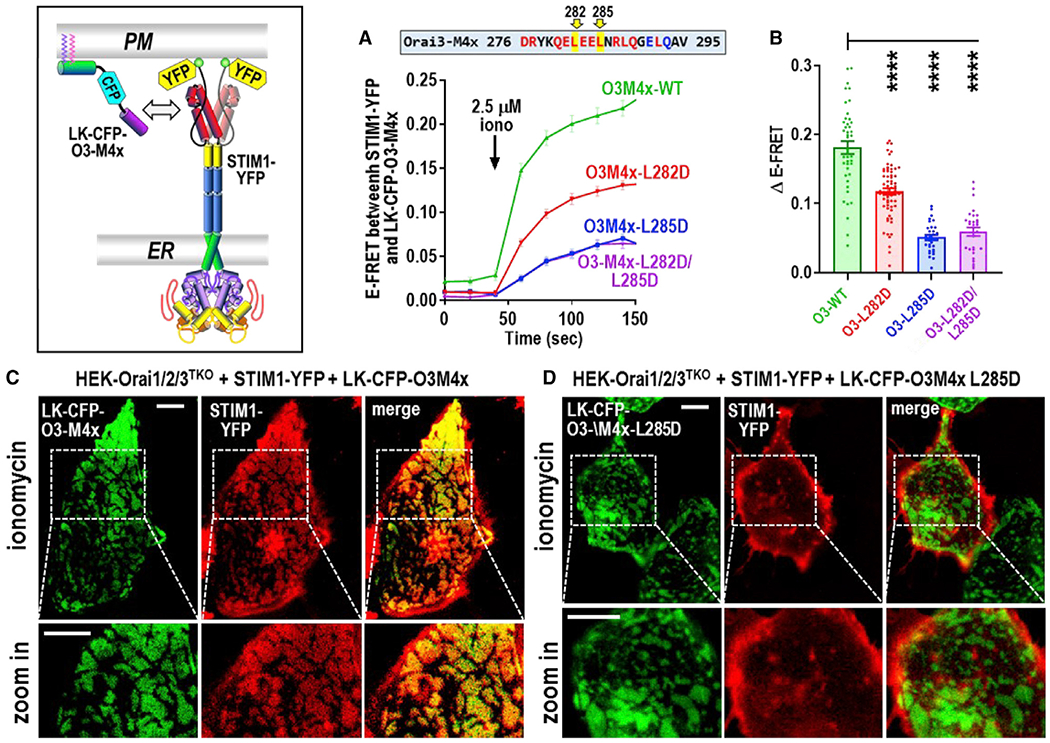
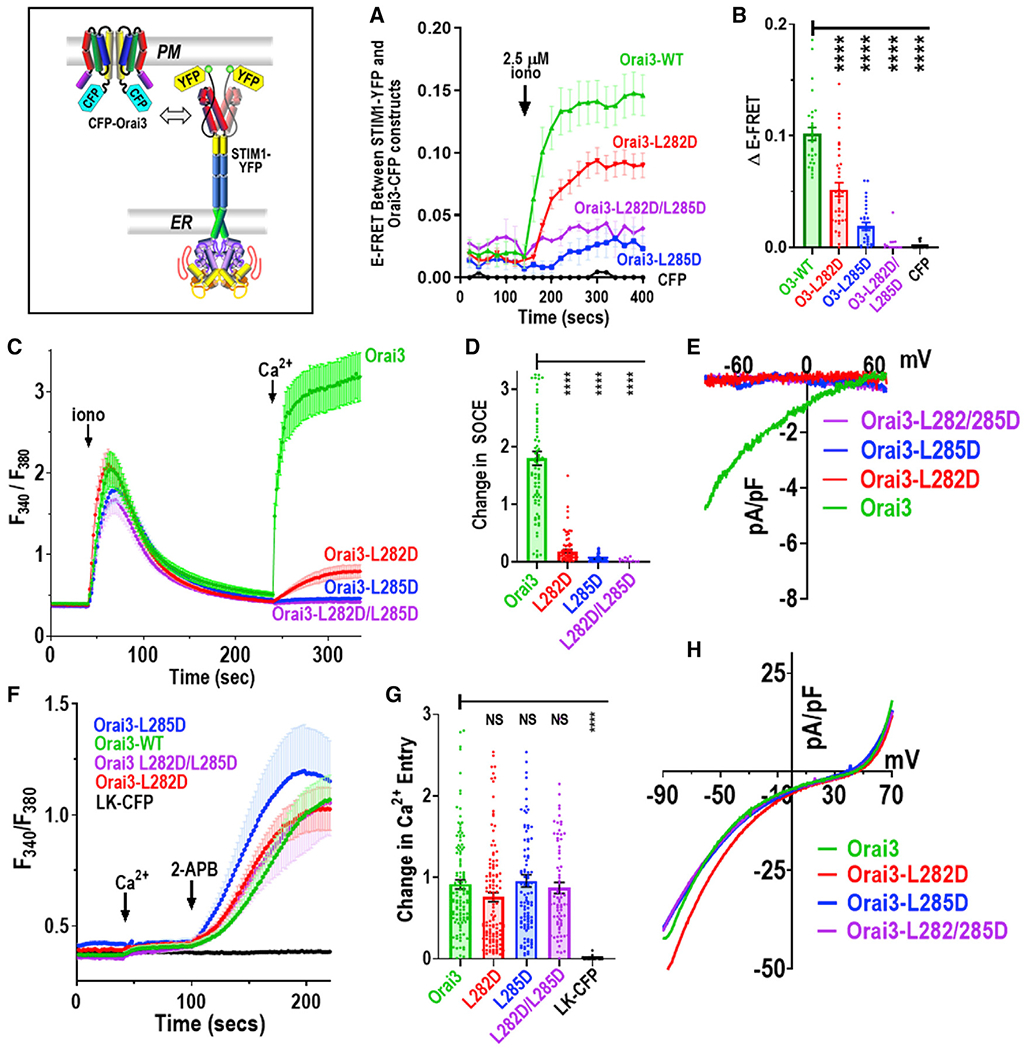
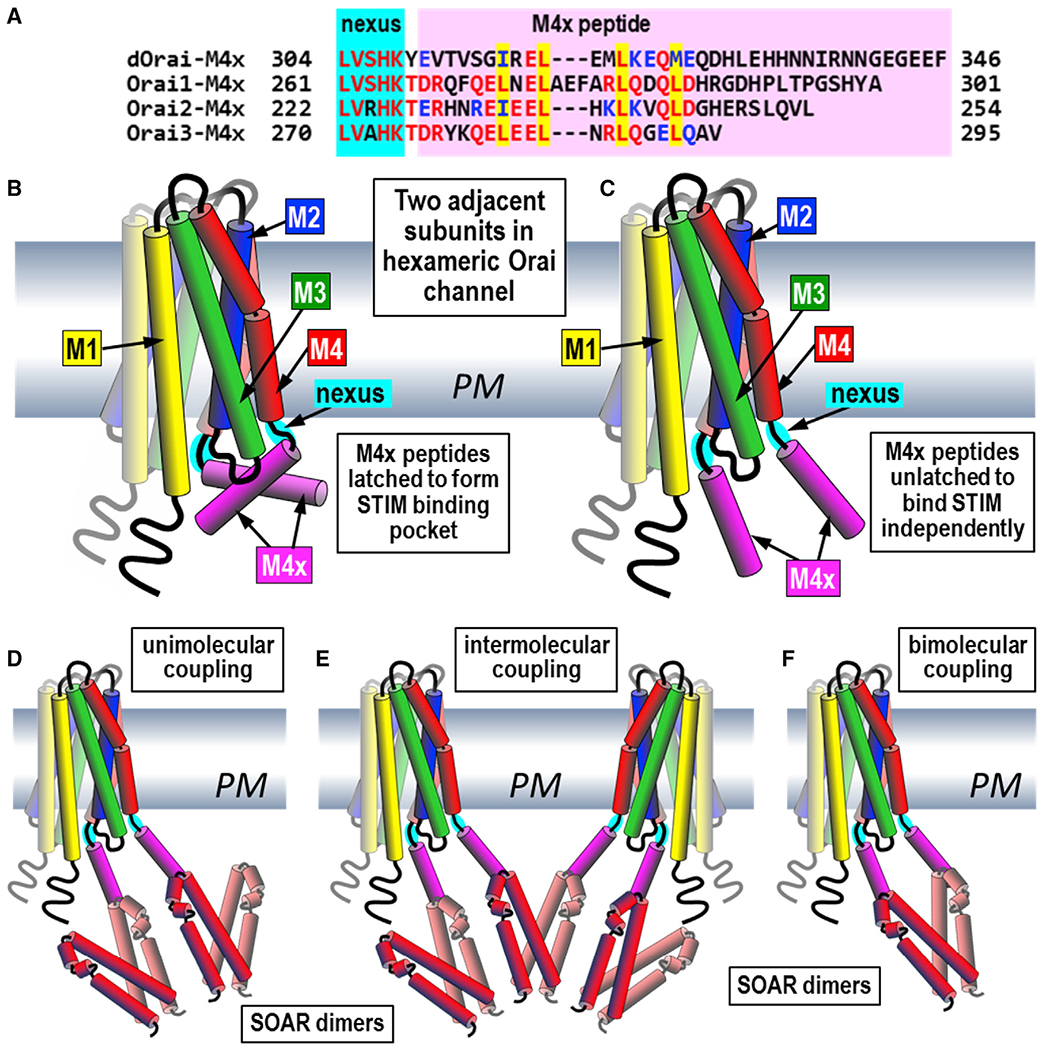
Junctional coupling between endoplasmic reticulum (ER) Ca2+-sensor STIM proteins and plasma membrane (PM) Orai channels mediates Ca2+ signals in most cells. We reveal that PM-tethered, fluorescently tagged C-terminal M4x (fourth transmembrane helix contains a cytoplasmic C-terminal extension) peptides from Orai channels undergo a Leu-specific signature of direct interaction with the STIM1 Orai-activating region (SOAR), exactly mimicking STIM1 binding to gate Orai channels. The 20-amino-acid Orai3-M4x peptide associates avidly with STIM1 within ER-PM junctions, functions to competitively block native Ca2+ signals, and mediates a key modification of STIM-Orai coupling induced by 2-aminoethoxydiphenyl borate. By blocking STIM-Orai coupling, the Orai3-M4x peptide reveals the critical role of Orai channels in driving Ca2+ oscillatory signals and transcriptional control through NFAT. The M4x peptides interact independently with SOAR dimers consistent with unimolecular coupling between Orai subunits and STIM1 dimers. We reveal the critical role of M4x helices in defining the coupling interface between STIM and Orai proteins to mediate store-operated Ca2+ signals.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



Comment in
-
Commentary to Baraniak et al. "Orai channel C-terminal peptides are key modulators of STIM-Orai coupling and calcium signal generation" published in cell reports 35: 109322.Cell Calcium. 2021 Sep;98:102455. doi: 10.1016/j.ceca.2021.102455. Epub 2021 Aug 18. Cell Calcium. 2021. PMID: 34433107 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
