

Screening and validation of genome-edited animals
- PMID: 34192966
- PMCID: PMC9008557
- DOI: 10.1177/00236772211016922
Screening and validation of genome-edited animals
Abstract
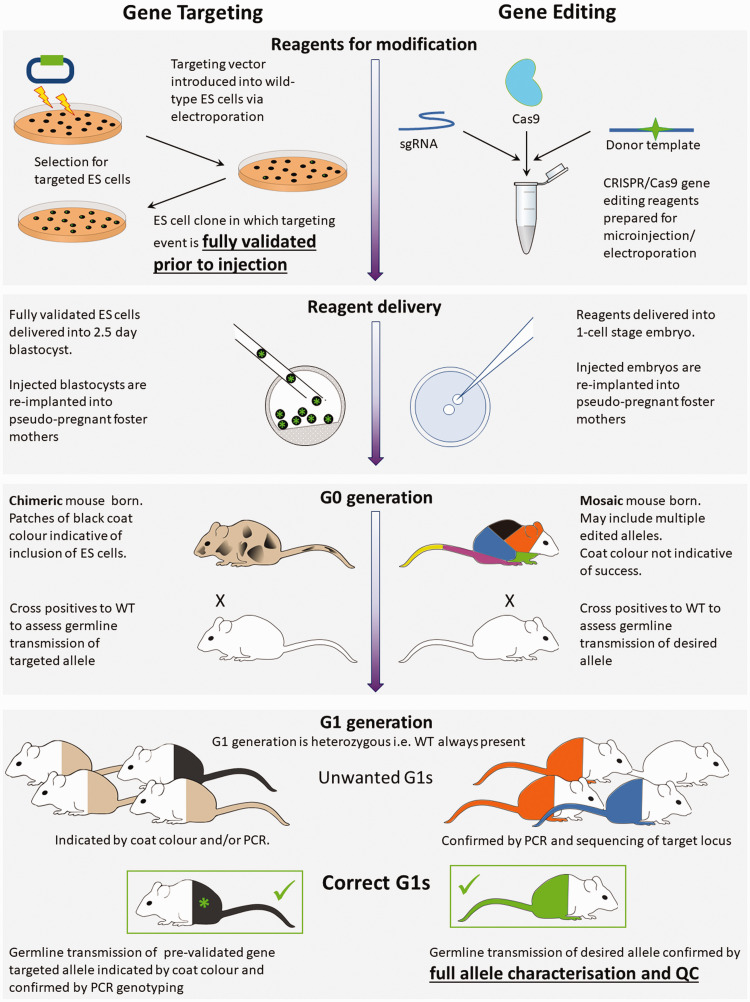
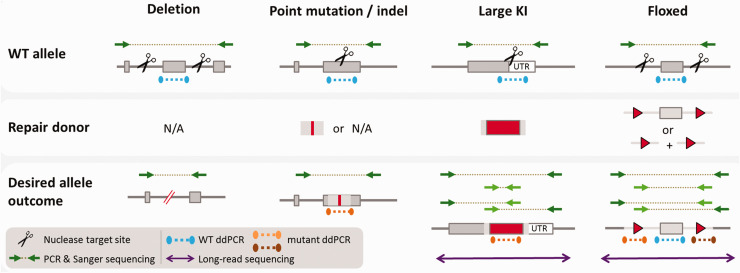
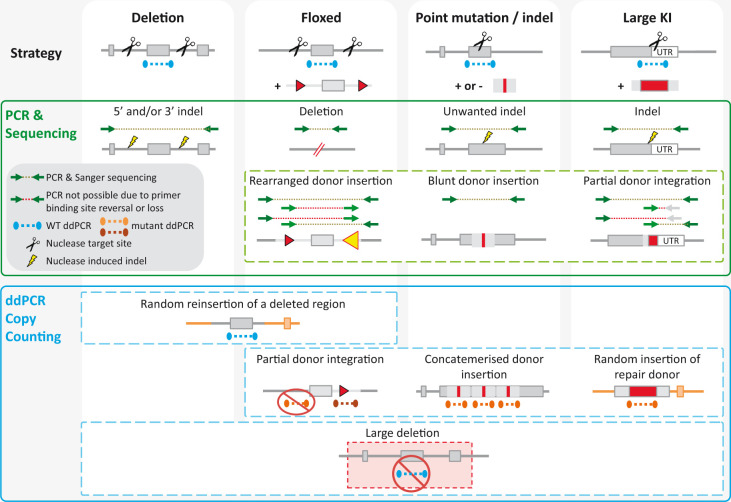
The emergence of an array of genome-editing tools in recent years has facilitated the introduction of genetic modifications directly into the embryo, increasing the ease, efficiency and catalogue of alleles accessible to researchers across a range of species. Bypassing the requirement for a selection cassette and resulting in a broad range of outcomes besides the desired allele, genome editing has altered the allele validation process both temporally and technically. Whereas traditional gene targeting relies upon selection and allows allele validation at the embryonic stem cell modification stage, screening for the presence of the intended allele now occurs in the (frequently mosaic) founder animals. Final confirmation of the edited allele can only take place at the subsequent G1 generation and the validation strategy must differentiate the desired allele from a range of unintended outcomes. Here we present some of the challenges posed by gene editing, strategies for validation and considerations for animal colony management.
L'émergence ces dernières années d'une gamme d'outils d'édition du génome a facilité l'introduction de modifications génétiques directement dans l'embryon, augmentant ainsi la facilité, l'efficacité et le catalogue des allèles accessibles aux chercheurs dans une gamme d'espèces. En contournant l'exigence d'une cassette de sélection et en permettant une large gamme de résultats outre l'allèle désiré, l'édition du génome a modifié le processus de validation d'allèle à la fois temporellement et techniquement. Alors que le ciblage traditionnel des gènes repose sur la sélection et permet une validation d'allèle au stade de la modification des cellules souches embryonnaires, le dépistage de la présence de l'allèle prévu se produit maintenant chez les animaux fondateurs (qui sont souvent mosaïques). La confirmation finale de l'allèle modifié ne peut avoir lieu qu'à la génération suivante (G1) et la stratégie de validation doit différencier l'allèle désiré d'une gamme de résultats inattendus. Nous présentons ici quelques-uns des défis posés par l'édition génétique, les stratégies de validation et les considérations pour la gestion des colonies animales.
Die Entwicklung einer Reihe von Tools zur Genom-Editierung hat in den letzten Jahren die Einführung von genetischen Modifikationen direkt in den Embryo begünstigt und die Einfachheit, Effizienz und den Katalog der Allele, die den Forschern über verschiedene Arten zugänglich sind, erhöht. Das Genome Editing umgeht die Notwendigkeit einer Selektionskassette und führt zu einer breiten Palette von Ergebnissen neben dem gewünschten Allel. Dadurch hat sich der Prozess der Allel-Validierung sowohl zeitlich als auch technisch verändert. Während das traditionelle Gen-Targeting auf Selektion beruht und eine Allel-Validierung im Stadium der ES-Zell-Modifikation ermöglicht, erfolgt das Screening auf das Vorhandensein des gewünschten Allels nun in den (häufig mosaischen) Gründertieren. Die endgültige Bestätigung des editierten Allels kann erst in der nachfolgenden G1-Generation erfolgen, und die Validierungsstrategie muss das gewünschte Allel von einer Reihe von unbeabsichtigten Ergebnissen unterscheiden. Hier stellen wir einige der Herausforderungen im Zusammenhang mit Gen-Editierung, Strategien zur Validierung und Überlegungen zum Management von Tierkolonien vor.
La emergencia en los últimos años de una serie de herramientas de modificación del genoma ha facilitado la introducción de modificaciones genéticas directamente en el embrión, aumentando así la facilidad, la eficiencia y las opciones de alelos accesibles para los investigadores en una gran variedad de especies. Al poder saltarnos la necesidad de un grupo de selección cerrado y con la posibilidad de una amplia gama de resultados además del alelo deseado, la modificación del genoma ha alterado el proceso de validación de alelos tanto temporal como técnicamente. Mientras que la selección tradicional de genes depende de la selección y permite la validación de alelos en la fase de modificación de células madre, hacer un cribado para identificar la presencia del alelo deseado ahora ocurre con los animales fundadores (normalmente mosaico). La confirmación final del alelo modificado solo puede hacerse en la siguiente generación G1 y la estrategia de validación debe diferenciar el alelo requerido de una serie de resultados no intencionados. En este estudio presentamos algunos de los retos que supone la edición genética, así como estrategias de validación y consideraciones para la gestión de colonias de animales.
Keywords: Animal model; GM; PCR; Quality assurance / control.
Conflict of interest statement
Figures

References
-
- Robertson E, Bradley A, Kuehn M, et al. Germ-line transmission of genes introduced into cultured pluripotential cells by retroviral vector. Nature 1986; 323: 445–448. - PubMed
-
- Doetschman T, Gregg RG, Maeda N, et al. Targeted correction of a mutant HPRT gene in mouse embryonic stem cells. Nature 1987; 330: 576–578. - PubMed
-
- Valenzuela DM, Murphy AJ, Frendewey D, et al. High-throughput engineering of the mouse genome coupled with high-resolution expression analysis. Nat Biotechnol 2003; 21: 652–659. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
