

Recapitulation of selective nuclear import and export with a perfectly repeated 12mer GLFG peptide
- PMID: 34193851
- PMCID: PMC8245513
- DOI: 10.1038/s41467-021-24292-5
Recapitulation of selective nuclear import and export with a perfectly repeated 12mer GLFG peptide
Abstract
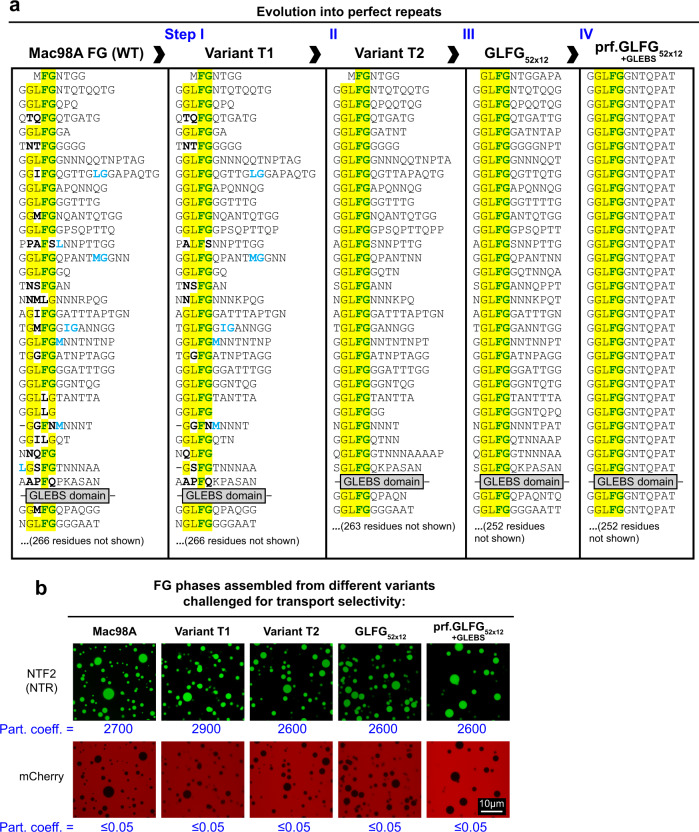
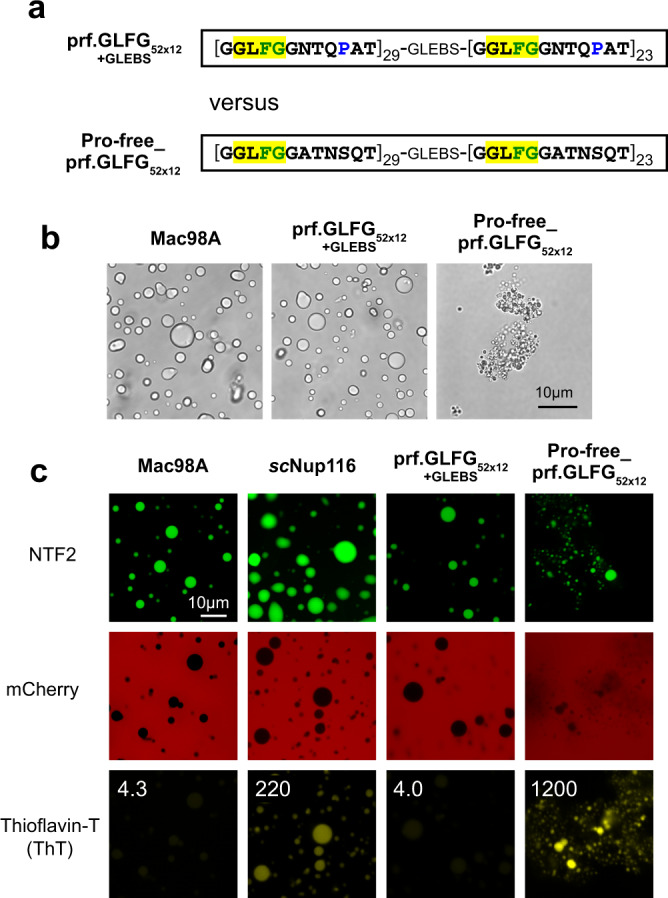
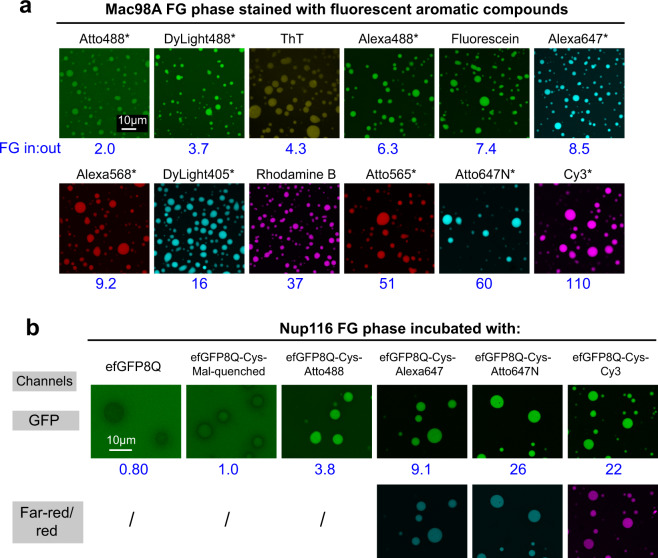
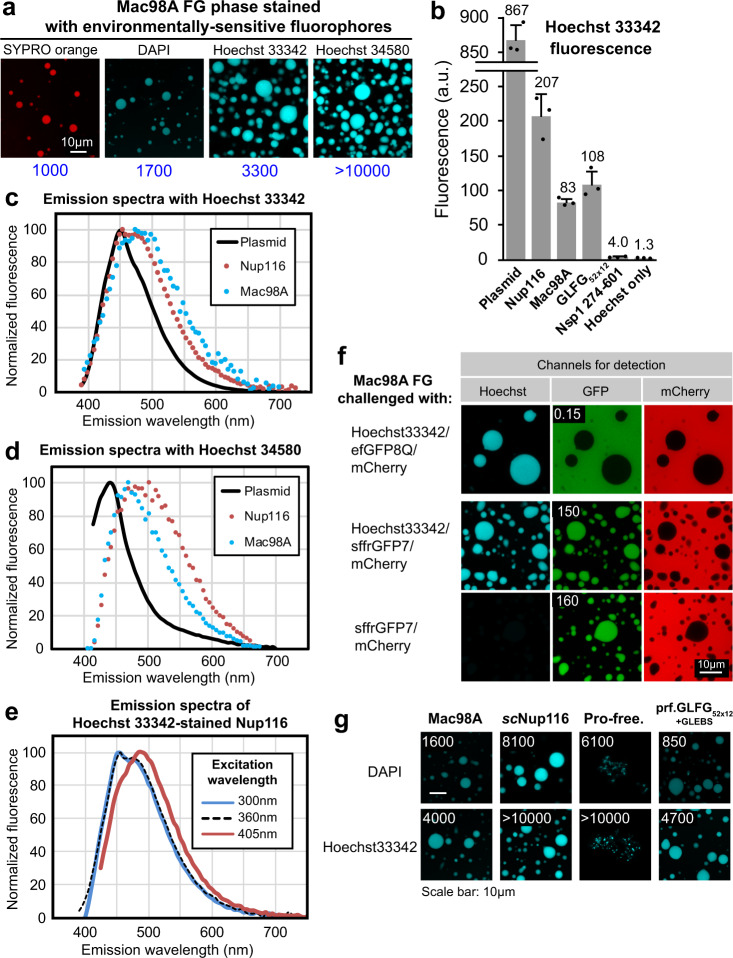
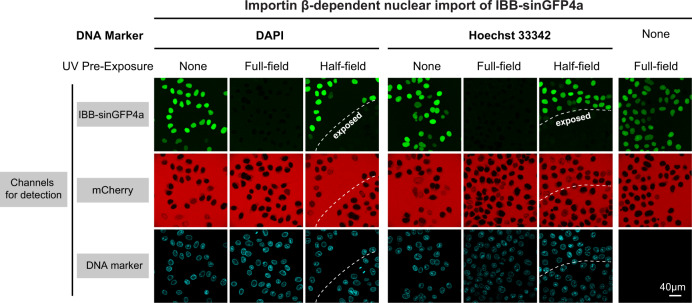
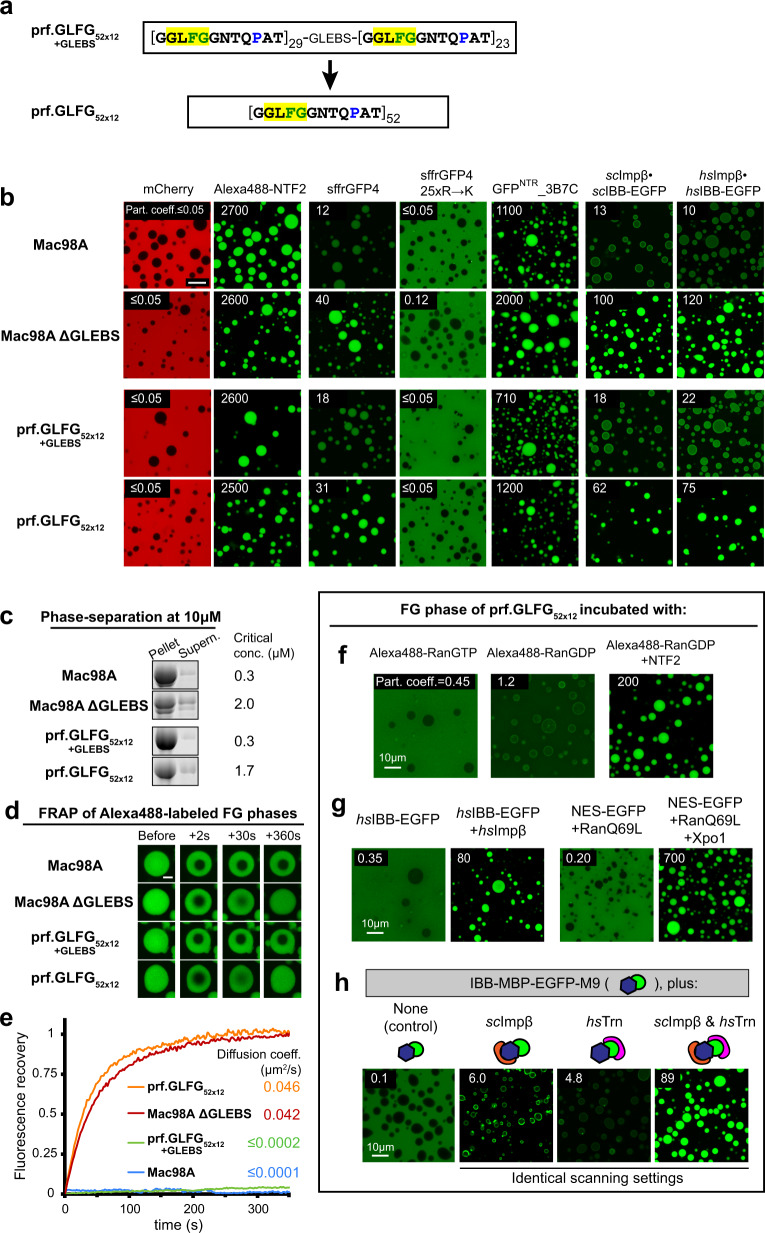
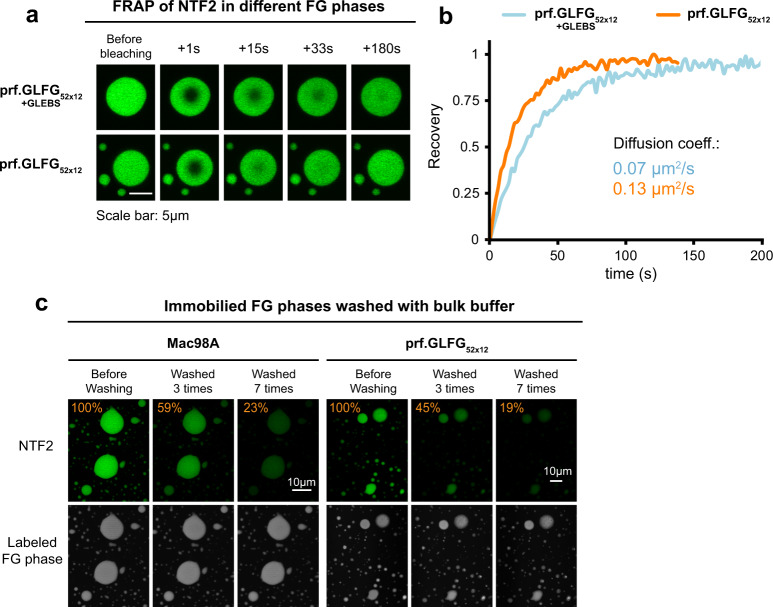
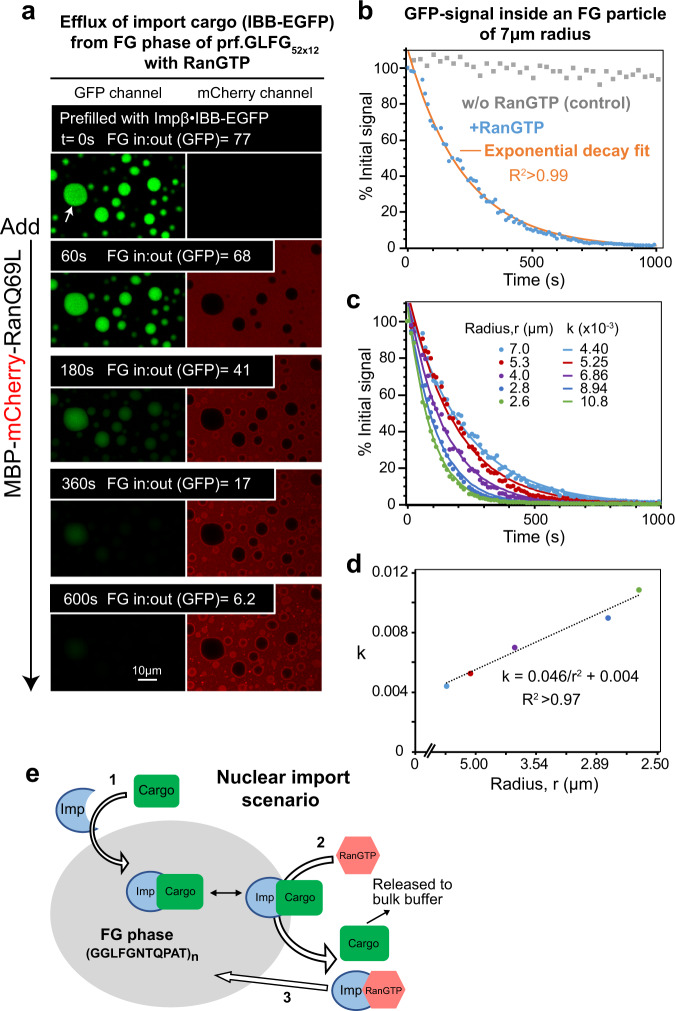
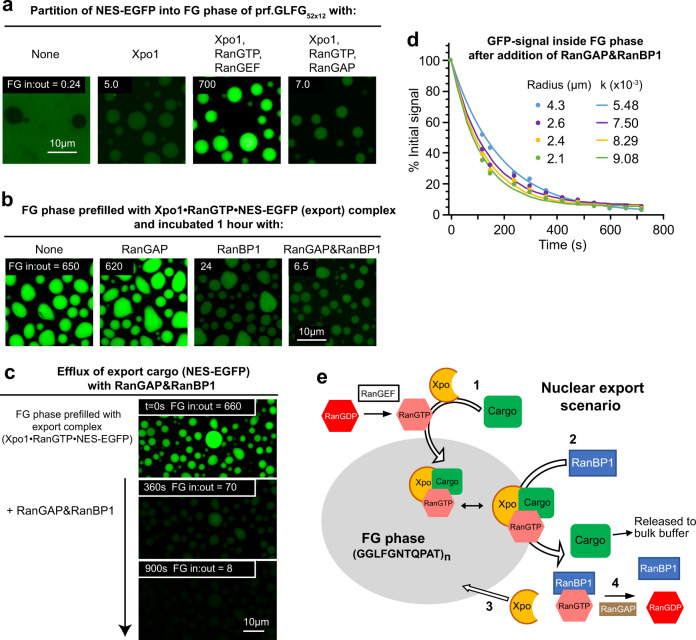
The permeability barrier of nuclear pore complexes (NPCs) controls nucleocytoplasmic transport. It retains inert macromolecules while allowing facilitated passage of importins and exportins, which in turn shuttle cargo into or out of cell nuclei. The barrier can be described as a condensed phase assembled from cohesive FG repeat domains. NPCs contain several distinct FG domains, each comprising variable repeats. Nevertheless, we now found that sequence heterogeneity is no fundamental requirement for barrier function. Instead, we succeeded in engineering a perfectly repeated 12mer GLFG peptide that self-assembles into a barrier of exquisite transport selectivity and fast transport kinetics. This barrier recapitulates RanGTPase-controlled importin- and exportin-mediated cargo transport and thus represents an ultimately simplified experimental model system. An alternative proline-free sequence forms an amyloid FG phase. Finally, we discovered that FG phases stain bright with 'DNA-specific' DAPI/ Hoechst probes, and that such dyes allow for a photo-induced block of nuclear transport.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
