Sleep/wake calcium dynamics, respiratory function, and ROS production in cardiac mitochondria
- PMID: 34194831
- PMCID: PMC8240107
- DOI: 10.1016/j.jare.2021.01.006
Sleep/wake calcium dynamics, respiratory function, and ROS production in cardiac mitochondria
Abstract
Introduction: Incidents of myocardial infarction and sudden cardiac arrest vary with time of the day, but the mechanism for this effect is not clear. We hypothesized that diurnal changes in the ability of cardiac mitochondria to control calcium homeostasis dictate vulnerability to cardiovascular events.
Objectives: Here we investigate mitochondrial calcium dynamics, respiratory function, and reactive oxygen species (ROS) production in mouse heart during different phases of wake versus sleep periods.
Methods: We assessed time-of-the-day dependence of calcium retention capacity of isolated heart mitochondria from young male C57BL6 mice. Rhythmicity of mitochondrial-dependent oxygen consumption, ROS production and transmembrane potential in homogenates were explored using the Oroboros O2k Station equipped with a fluorescence detection module. Changes in expression of essential clock and calcium dynamics genes/proteins were also determined at sleep versus wake time points.
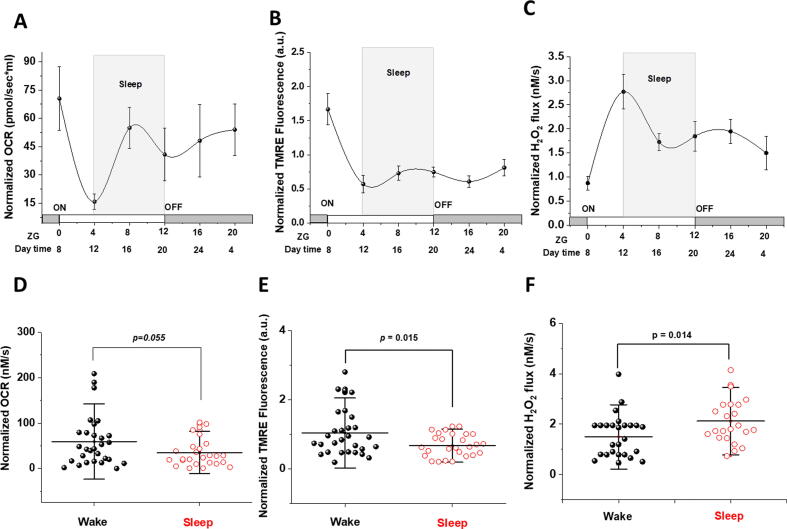
Results: Our results demonstrate that cardiac mitochondria exhibit higher calcium retention capacity and higher rates of calcium uptake during sleep period. This was associated with higher expression of clock gene Bmal1, lower expression of per2, greater expression of MICU1 gene (mitochondrial calcium uptake 1), and lower expression of the mitochondrial transition pore regulator gene cyclophilin D. Protein levels of mitochondrial calcium uniporter (MCU), MICU2, and sodium/calcium exchanger (NCLX) were also higher at sleep onset relative to wake period. While complex I and II-dependent oxygen utilization and transmembrane potential of cardiac mitochondria were lower during sleep, ROS production was increased presumably due to mitochondrial calcium sequestration.
Conclusions: Taken together, our results indicate that retaining mitochondrial calcium in the heart during sleep dissipates membrane potential, slows respiratory activities, and increases ROS levels, which may contribute to increased vulnerability to cardiac stress during sleep-wake transition. This pronounced daily oscillations in mitochondrial functions pertaining to stress vulnerability may at least in part explain diurnal prevalence of cardiac pathologies.
Keywords: Calcium dynamics; Clock genes; Diurnal; Heart; Hydrogen peroxide; Mitochondria function.
© 2021 The Authors. Published by Elsevier B.V. on behalf of Cairo University.
Conflict of interest statement
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: HHP has equity as a founder in CavoGene LifeSciences Holdings, LLC. All other authors have declared no conflict of interest.
Figures