

Minor Intron Splicing from Basic Science to Disease
- PMID: 34199764
- PMCID: PMC8199999
- DOI: 10.3390/ijms22116062
Minor Intron Splicing from Basic Science to Disease
Abstract
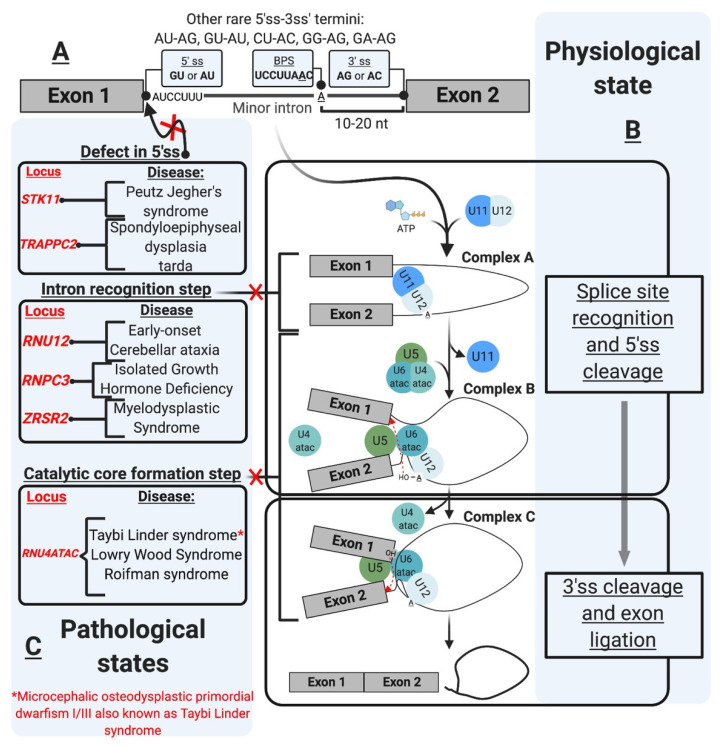
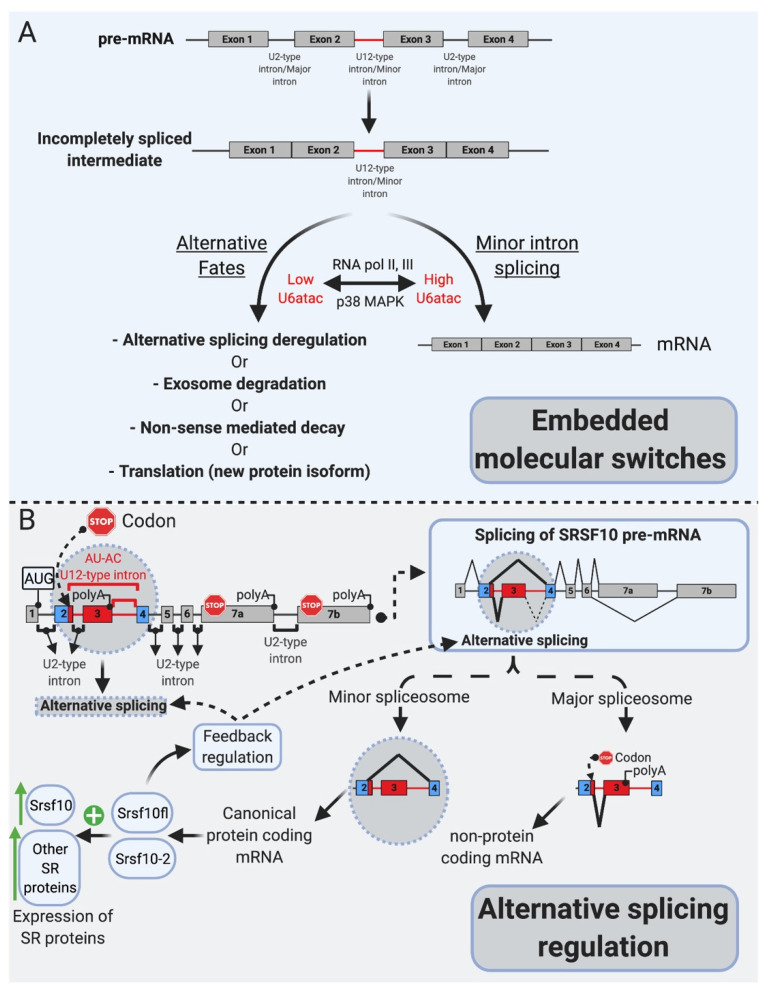
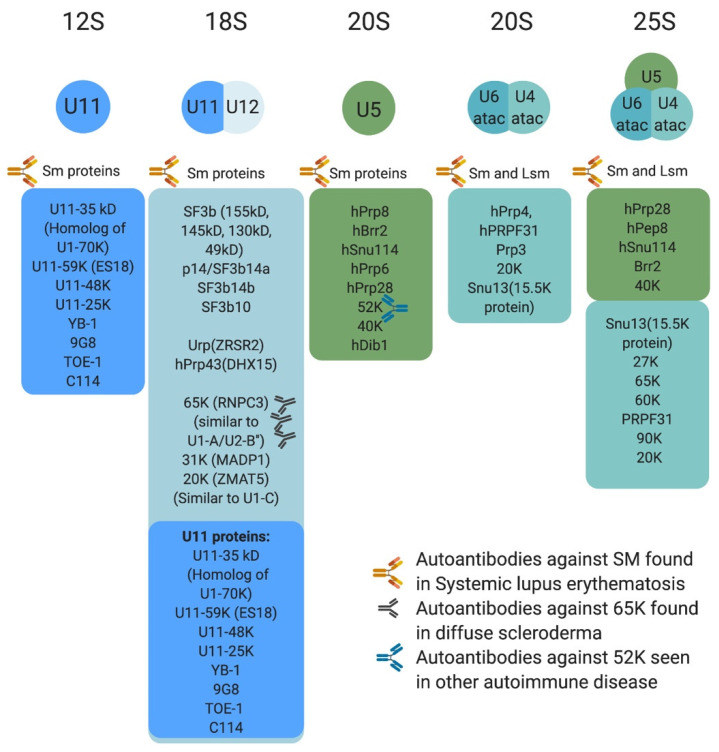
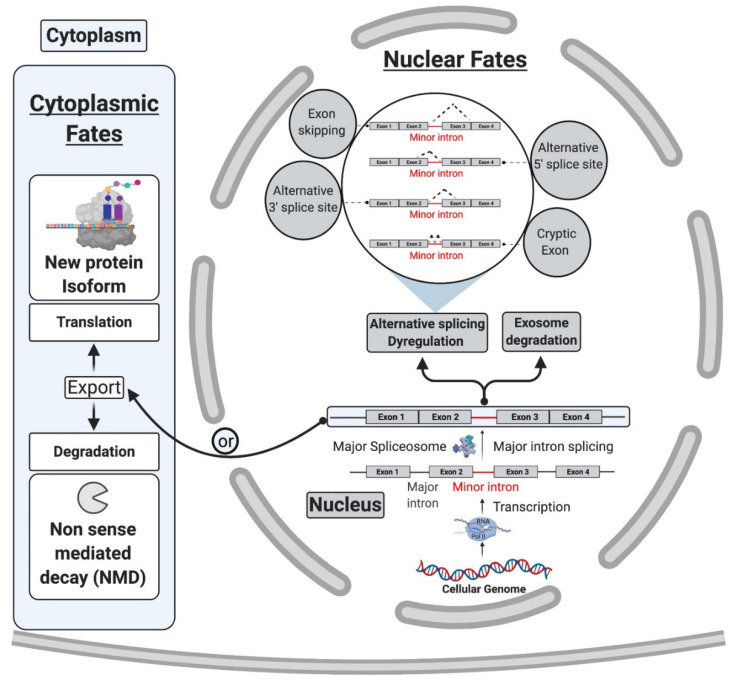
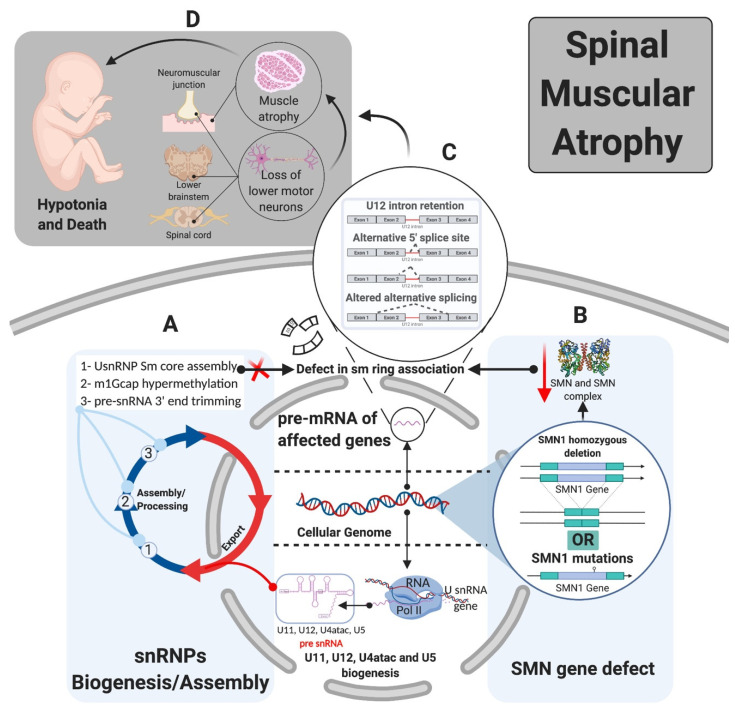
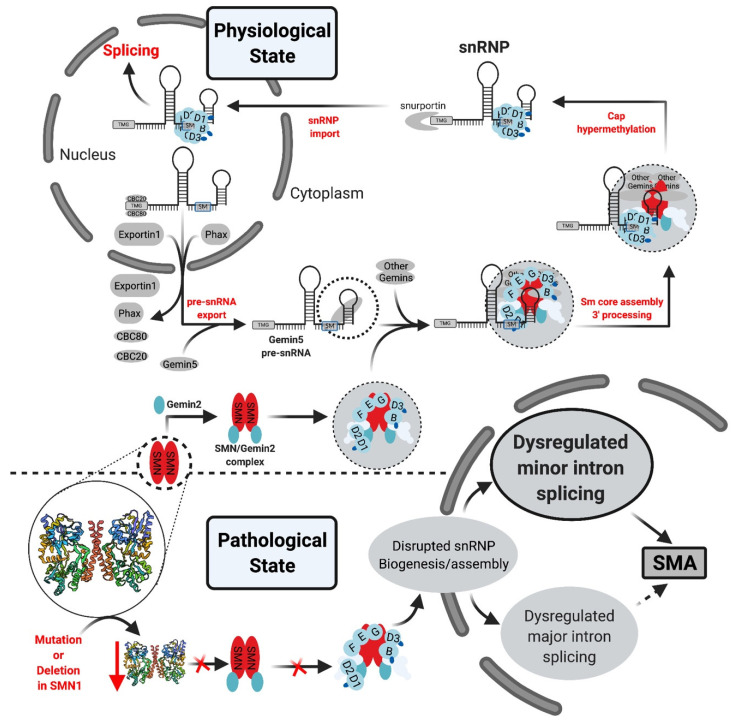
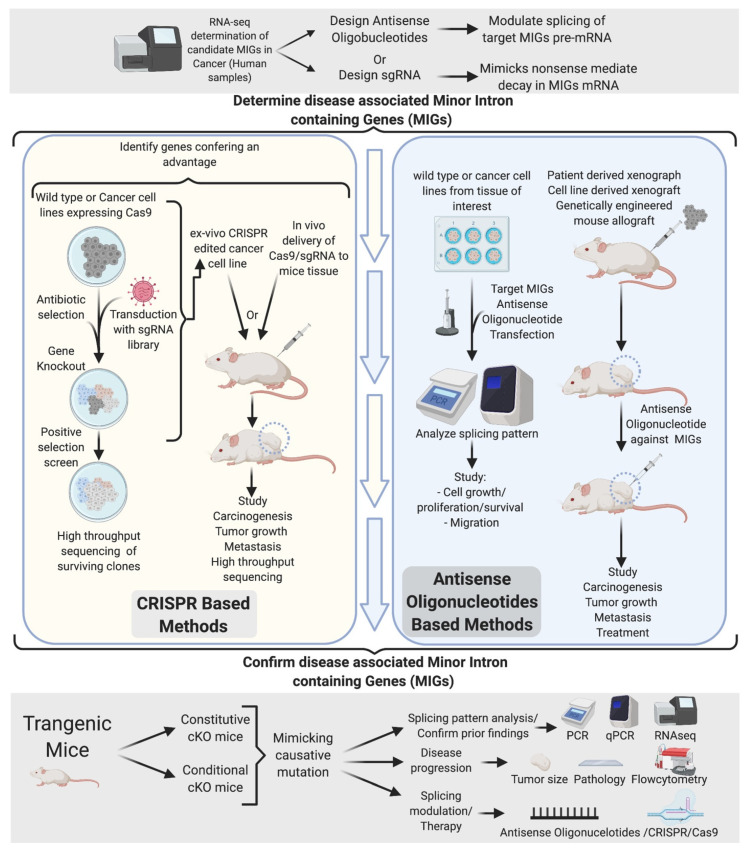
Pre-mRNA splicing is an essential step in gene expression and is catalyzed by two machineries in eukaryotes: the major (U2 type) and minor (U12 type) spliceosomes. While the majority of introns in humans are U2 type, less than 0.4% are U12 type, also known as minor introns (mi-INTs), and require a specialized spliceosome composed of U11, U12, U4atac, U5, and U6atac snRNPs. The high evolutionary conservation and apparent splicing inefficiency of U12 introns have set them apart from their major counterparts and led to speculations on the purpose for their existence. However, recent studies challenged the simple concept of mi-INTs splicing inefficiency due to low abundance of their spliceosome and confirmed their regulatory role in alternative splicing, significantly impacting the expression of their host genes. Additionally, a growing list of minor spliceosome-associated diseases with tissue-specific pathologies affirmed the importance of minor splicing as a key regulatory pathway, which when deregulated could lead to tissue-specific pathologies due to specific alterations in the expression of some minor-intron-containing genes. Consequently, uncovering how mi-INTs splicing is regulated in a tissue-specific manner would allow for better understanding of disease pathogenesis and pave the way for novel therapies, which we highlight in this review.
Keywords: RNA splicing; U12 introns; U2 introns; disease; minor introns; minor spliceosome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
