

Colombian Scorpion Centruroides margaritatus: Purification and Characterization of a Gamma Potassium Toxin with Full-Block Activity on the hERG1 Channel
- PMID: 34201318
- PMCID: PMC8273696
- DOI: 10.3390/toxins13060407
Colombian Scorpion Centruroides margaritatus: Purification and Characterization of a Gamma Potassium Toxin with Full-Block Activity on the hERG1 Channel
Abstract
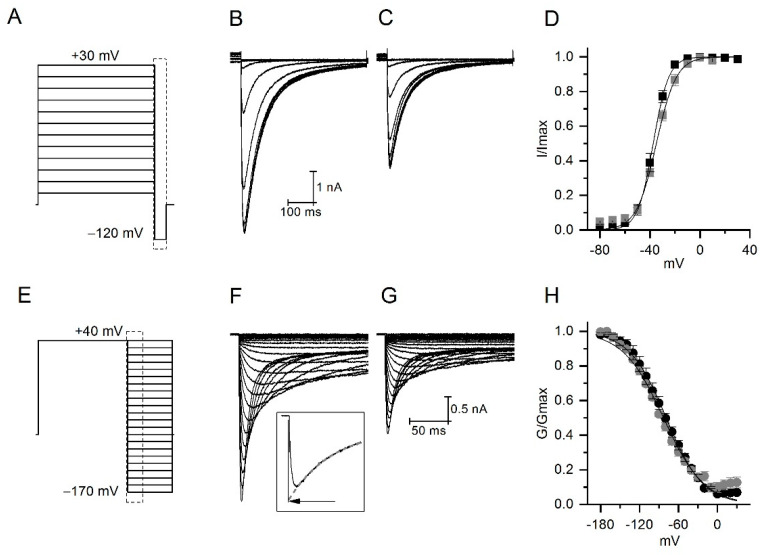
The Colombian scorpion Centruroides margaritatus produces a venom considered of low toxicity. Nevertheless, there are known cases of envenomation resulting in cardiovascular disorders, probably due to venom components that target ion channels. Among them, the humanether-à-go-go-Related gene (hERG1) potassium channels are critical for cardiac action potential repolarization and alteration in its functionality are associated with cardiac disorders. This work describes the purification and electrophysiological characterization of a Centruroides margaritatus venom component acting on hERG1 channels, the CmERG1 toxin. This novel peptide is composed of 42 amino acids with a MW of 4792.88 Da, folded by four disulfide bonds and it is classified as member number 10 of the γ-KTx1 toxin family. CmERG1 inhibits hERG1 currents with an IC50 of 3.4 ± 0.2 nM. Despite its 90.5% identity with toxin ɣ-KTx1.1, isolated from Centruroides noxius, CmERG1 completely blocks hERG1 current, suggesting a more stable plug of the hERG channel, compared to that formed by other ɣ-KTx.
Keywords: Centruroides margaritatus; CmERG1; CnERG1; ERG channel; ERG toxin; Electrophysiology.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








References
-
- De Armas L.F., Sarmiento D.L., Flórez E.L. Composición del género Centruroides marx, 1890 (Scorpiones buthidae) en Colombia, con la descripción de una nueva especie. Boletín Soc. Entomológica Aragonsa. 2012;50:105–114.
-
- Guerrero-Vargas J.A., Rodríguez Buitrago J.R., Ayerbe S., Flórez Daza E., Beltrán Vidal J.T. Scorpionism and Dangerous Species of ColombiaColombia. In: Gopalakrishnakone P., Possani L.D., F. Schwartz E., Rodríguez de la Vega R.C., editors. Scorpion Venoms. Springer; Dordrecht, The Netherlands: 2015. pp. 245–272. - DOI
Publication types
MeSH terms
Substances
Grants and funding
- IN202619/Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México
- PRONACE 303045.P/Consejo Nacional de Ciencia y Tecnología
- Movilidad Internacional CIAM/Vicerrectoría de Investigaciones, Universidad del Valle, Colombia
- Movilidad Internacional/Vicerrectoría de Investigaciones, Universidad del Cauca,Colombia
LinkOut - more resources
Full Text Sources
