

Heterochromatin Networks: Topology, Dynamics, and Function (a Working Hypothesis)
- PMID: 34201566
- PMCID: PMC8304199
- DOI: 10.3390/cells10071582
Heterochromatin Networks: Topology, Dynamics, and Function (a Working Hypothesis)
Abstract
Open systems can only exist by self-organization as pulsing structures exchanging matter and energy with the outer world. This review is an attempt to reveal the organizational principles of the heterochromatin supra-intra-chromosomal network in terms of nonlinear thermodynamics. The accessibility of the linear information of the genetic code is regulated by constitutive heterochromatin (CHR) creating the positional information in a system of coordinates. These features include scale-free splitting-fusing of CHR with the boundary constraints of the nucleolus and nuclear envelope. The analysis of both the literature and our own data suggests a radial-concentric network as the main structural organization principle of CHR regulating transcriptional pulsing. The dynamic CHR network is likely created together with nucleolus-associated chromatin domains, while the alveoli of this network, including springy splicing speckles, are the pulsing transcription hubs. CHR contributes to this regulation due to the silencing position variegation effect, stickiness, and flexible rigidity determined by the positioning of nucleosomes. The whole system acts in concert with the elastic nuclear actomyosin network which also emerges by self-organization during the transcriptional pulsing process. We hypothesize that the the transcriptional pulsing, in turn, adjusts its frequency/amplitudes specified by topologically associating domains to the replication timing code that determines epigenetic differentiation memory.
Keywords: chromatin organization; cytoskeleton; heterochromatin; networks; nucleolar boundary; physics of life; positional information; scale-free oscillations; transcriptional pulsing.
Conflict of interest statement
The authors declare no conflict of interests.
Figures


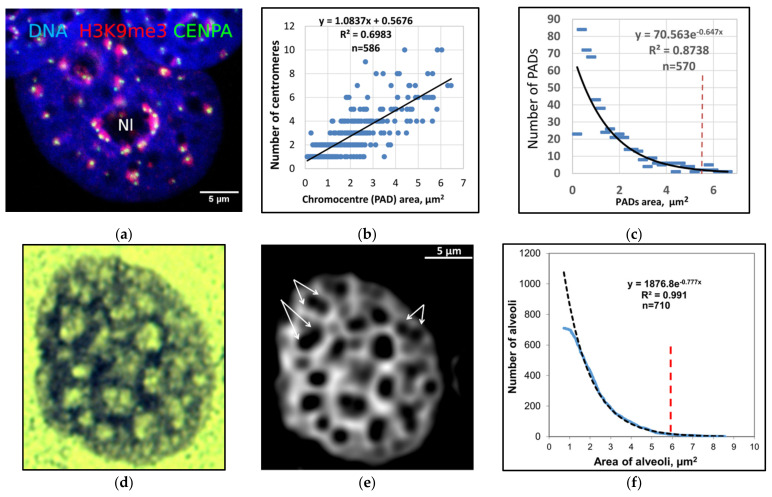
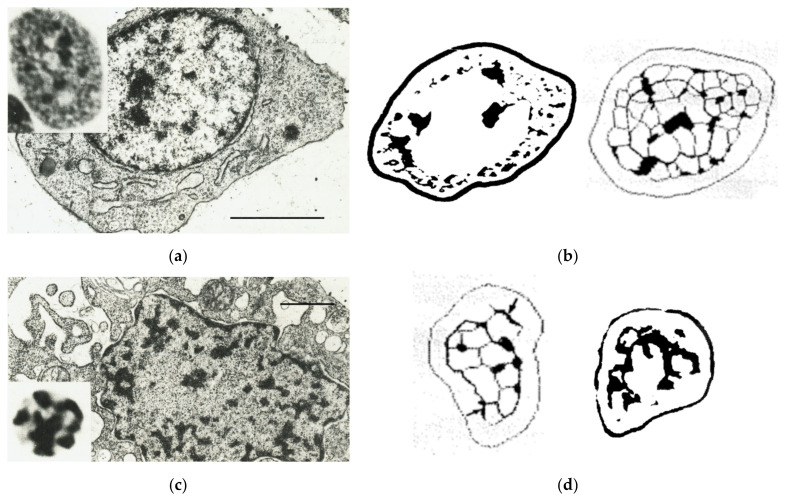
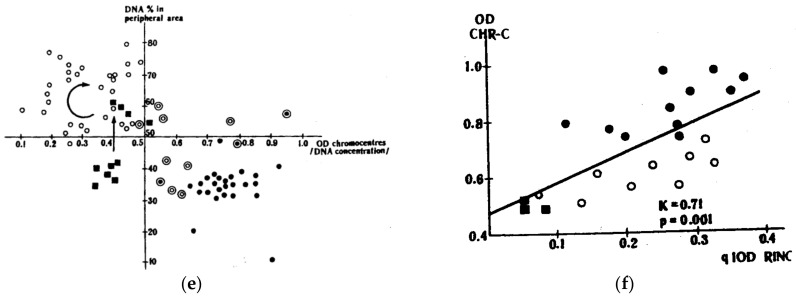
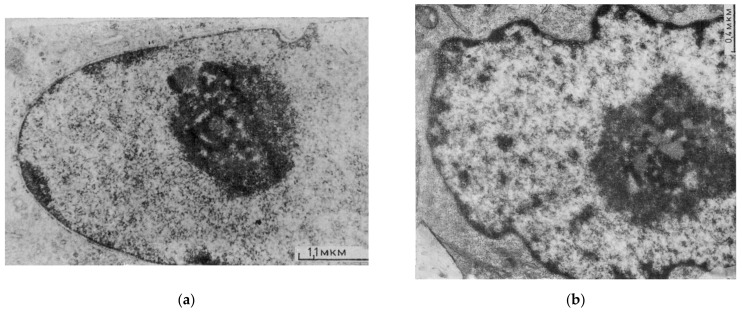
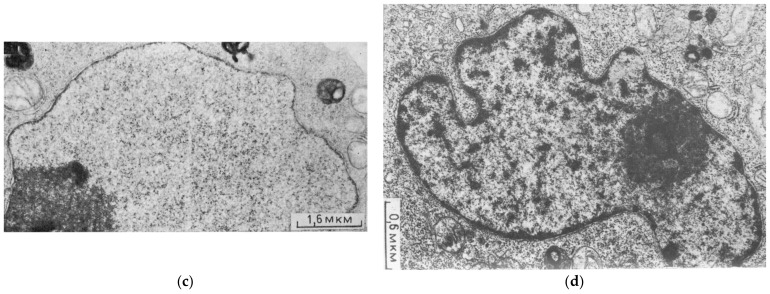
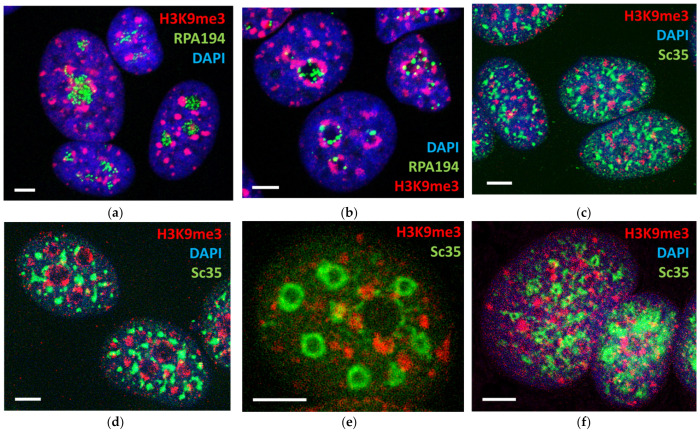
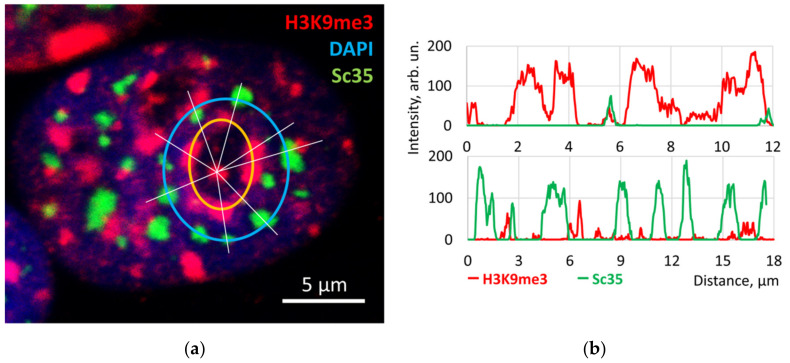
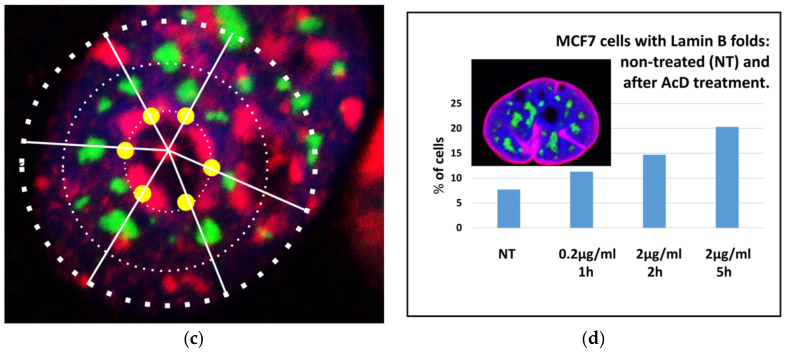

Similar articles
-
Close to the edge: Heterochromatin at the nucleolar and nuclear peripheries.Biochim Biophys Acta Gene Regul Mech. 2021 Jan;1864(1):194666. doi: 10.1016/j.bbagrm.2020.194666. Epub 2020 Dec 8. Biochim Biophys Acta Gene Regul Mech. 2021. PMID: 33307247 Free PMC article. Review.
-
Loss of nucleolar histone chaperone NPM1 triggers rearrangement of heterochromatin and synergizes with a deficiency in DNA methyltransferase DNMT3A to drive ribosomal DNA transcription.J Biol Chem. 2014 Dec 12;289(50):34601-19. doi: 10.1074/jbc.M114.569244. Epub 2014 Oct 27. J Biol Chem. 2014. PMID: 25349213 Free PMC article.
-
Nucleolar aggresomes mediate release of pericentric heterochromatin and nuclear destruction of genotoxically treated cancer cells.Nucleus. 2017 Mar 4;8(2):205-221. doi: 10.1080/19491034.2017.1279775. Epub 2017 Jan 9. Nucleus. 2017. PMID: 28068183 Free PMC article.
-
Distinct features of nucleolus-associated domains in mouse embryonic stem cells.Chromosoma. 2020 Jun;129(2):121-139. doi: 10.1007/s00412-020-00734-9. Epub 2020 Mar 26. Chromosoma. 2020. PMID: 32219510 Free PMC article.
-
Nuclear Architecture of Mouse Spermatocytes: Chromosome Topology, Heterochromatin, and Nucleolus.Cytogenet Genome Res. 2017;151(2):61-71. doi: 10.1159/000460811. Epub 2017 May 12. Cytogenet Genome Res. 2017. PMID: 28494440 Review.
Cited by
-
Advanced image-free analysis of the nano-organization of chromatin and other biomolecules by Single Molecule Localization Microscopy (SMLM).Comput Struct Biotechnol J. 2023 Mar 9;21:2018-2034. doi: 10.1016/j.csbj.2023.03.009. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 36968017 Free PMC article.
-
Polyploidy and Myc Proto-Oncogenes Promote Stress Adaptation via Epigenetic Plasticity and Gene Regulatory Network Rewiring.Int J Mol Sci. 2022 Aug 26;23(17):9691. doi: 10.3390/ijms23179691. Int J Mol Sci. 2022. PMID: 36077092 Free PMC article. Review.
-
Nuclear Fragility in Radiation-Induced Senescence: Blebs and Tubes Visualized by 3D Electron Microscopy.Cells. 2022 Jan 13;11(2):273. doi: 10.3390/cells11020273. Cells. 2022. PMID: 35053389 Free PMC article.
-
Transcriptome-Wide Insights: Neonatal Lactose Intolerance Promotes Telomere Damage, Senescence, and Cardiomyopathy in Adult Rat Heart.Int J Mol Sci. 2025 Feb 13;26(4):1584. doi: 10.3390/ijms26041584. Int J Mol Sci. 2025. PMID: 40004050 Free PMC article.
-
Spatial-Temporal Genome Regulation in Stress-Response and Cell-Fate Change.Int J Mol Sci. 2023 Jan 31;24(3):2658. doi: 10.3390/ijms24032658. Int J Mol Sci. 2023. PMID: 36769000 Free PMC article. Review.
References
-
- The Nobel Prize in Chemistry. [(accessed on 28 May 2021)];1977 Available online: https://www.nobelprize.org/prizes/chemistry/1977/prigogine/lecture/
-
- Schroedinger E. What Is Life? The Physical Aspect of the Living Cell, Etc. Cambridge University Press; Cambridge, UK: 1944.
-
- Knoch T.A. Chromatin and Epigenetics. IntechOpen; London, UK: 2020. A consistent systems mechanics model of the 3D architecture and dynamics of genomes.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
